bio.wikisort.org - Животные
Сина́псиды[1][2] (также употребляется синапси́ды[3]) (лат. Synapsida) — клада амниот, включающая млекопитающих и всех вымерших животных, более близких к млекопитающим, чем к завропсидам (рептилиям, включая птиц)[4]. Синапсиды легко отличаются от других амниот по височному отверстию, отверстию в нижней части черепной крыши за каждым глазом, образующим под ними костные дуги, что объясняет их название[5]. Возникли в среднем или раннем карбоне, около 310—330 млн лет назад.
| Синапсиды | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: Cotylorhynchus romeri, Dimetrodon grandis; | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Класс: Синапсиды |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Synapsida Osborn, 1903 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 318 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
На протяжении большей части XX века синапсиды считались одним из подклассов класса пресмыкающихся, не включающим млекопитающих. Так, в монографии Р. Кэрролла синапсиды выступали именно как подкласс класса пресмыкающиеся (Reptilia), включавший 2 отряда: пеликозавров (Pelycosauria) и терапсид (Therapsida)[6]. Позднее широкое распространение представлений, характерных для филогенетической систематики, привело к пересмотру классификации синапсид; в конце XX века они были выведены из состава класса пресмыкающихся[7][8].
По современным представлениям, синапсиды вместе с сестринской группой завропсид (рептилий) образуют кладу амниот — крупнейшую по объёму группу в составе надкласса четвероногих[9][10]. Поскольку в своём традиционном составе синапсиды являются парафилетической по отношению к млекопитающим группой, в научном сообществе сложился консенсус, согласно которому млекопитающие образуют подгруппу синапсид[11][7][8][lower-alpha 1]. Всех синапсид, за исключением млекопитающих, называют немаммальными синапсидами.
История изучения
Первые открытия


История изучения синапсид восходит к 1838 году. В этом году российский зоолог С. С. Куторга, исследуя находки, сделанные в 1770-е годы русскими горными инженерами в медистых песчаниках на западных склонах Уральских гор, дал первые научные описания ископаемых остатков синапсид, выделив виды Brithopus priscus, Orthopus primaevus и Syodon biarmicum. Эти виды учёный считал примитивными млекопитающими; по современным представлениям они относятся к группе дейноцефалов[13][14][15]. В 1841 году другой российский зоолог, Г. И. Фишер фон Вальдгейм, в ходе изучения ископаемых остатков из того же региона описал вид Rhopalodon wangenheimi (также ныне относимый к дейноцефалам)[16][17].
В 1845 году английский зоолог Ричард Оуэн, изучая находки, сделанные в регионе Кару на юге Африки Э. Бейном, описал вид Dicynodon lacerticeps — первого представителя группы дицинодонтов, причём отнёс данный вид к рептилиям[18]. В 1854 году американский зоолог Джозеф Лейди описал по найденной на канадском острове Принца Эдуарда части верхней челюсти вид Bathygnathus borealis (в 2015 году переклассифицирован как Dimetrodon borealis[19]) — первого пеликозавра[20]. Постепенно появлялись и новые находки; обобщая результаты первых исследований, Р. Оуэн установил в 1859 году отряд рептилий Anomodontia, а в 1876 году выделил из него отдельный отряд Theriodontia (включавший формы с многочисленными хорошо дифференцированными зубами). В 1878 году американский палеонтолог Эдвард Коп ввёл отряд Theromorpha с подразделением его на подотряды Pelycosauria и Anomodontia (последние, в свою очередь, включали группы Dicynodontia и Theriodontia), указав при этом на систематическую близость Theromorpha и млекопитающих[21][22]. Вскоре Коп, однако, расширил объём Theromorpha, включив в него также котилозавров[23]; в то же время выделенное им в 1884 году семейство Tritylodontidae он причислил не к Theromorpha, а к млекопитающим (лишь в 1956 году немецкий палеонтолог Вальтер Кюне отнёс это семейство к цинодонтам)[24].
Конец XIX — начало XX века

В течение XIX и начала XX века было сделано много находок ископаемых остатков синапсид — в основном в трёх удалённых друг от друга регионах: Южной Африке, Северной Америке и Восточной Европе (положение принципиально не изменилось и позднее: хотя остатки синапсид находили в самых различных местах земного шара, ведущими источниками сведений о синапсидной фауне оставались именно три упомянутых региона)[25]. Продолжалось — усилиями таких палеонтологов, как Р. Оуэн, Г. Сили, Р. Брум, Д. Уотсон — изучение региона Кару, в результате чего были описаны такие известные представители терапсид, как горгонопс, тритилодон, тринаксодон, диадемодон, трираходон, циногнат, каннемейерия, баурия, мосхопс, трителедон, арктопс[26][27]. Со 2-й половины 1870-х годов начались результативные поиски ископаемых остатков синапсид в Северной Америке: Э. Коп описал виды пеликозавров Clepsydrops collettii (1875) по находкам в Иллинойсе и Dimetrodon limbatus (1878) по находкам в Техасе, а оппонент Копа О. Ч. Марш в том же 1878 году по ископаемым остаткам из Нью-Мексико описал виды пеликозавров Sphenacodon ferox и Ophiacodon mirus[28]. Важнейшим этапом палеонтологического исследования Восточной Европы стали ежегодные экспедиции В. П. Амалицкого в бассейн Северной Двины (1895—1914 годы), в ходе которых были описаны, в частности, иностранцевия, двиния, венюковия[29][30].

Американский палеонтолог Генри Осборн в 1903 году ввёл деление класса Reptilia на два подкласса: Diapsida и Synapsida. Объём подкласса синапсид в трактовке Осборна, правда, существенно отличался от принимаемого ныне, так как Осборн включил в него наряду с надотрядом Anomodontia также котилозавров, черепах и завроптеригий (т. e. группы с анапсидным и эвриапсидным строением черепа), а отряд Pelycosauria отнёс к диапсидам (считая предложенное Копом их объединение с Anomodontia в одном таксоне и сам термин Theromorpha неправомерными). Другие палеонтологи ввели в эту классификацию уточнения, но само название Synapsida удержалось и со временем почти полностью вытеснило название Theromorpha[21][31].
В результате «лишние» группы были выведены из объёма Synapsida, и уже в 1925 году в фундаментальной монографии Сэмюэла Уиллистона «The Osteology of the Reptiles» данный таксон предстаёт — как и в большинстве последующих работ XX века — как подкласс, состоящий из двух отрядов: Pelycosauria Cope, 1878 (с более примитивными формами) и Therapsida Broom, 1905 (к которому были отнесёны более продвинутые формы). Сам Уиллистон, впрочем, первый из этих отрядов именовал Theromorpha, а объём Pelycosauria трактовал более узко; в 1940 году Альфред Ромер и Ллевеллин Прайс в своём исследовании «Review of the Pelycosauria»[32] приняли для данного отряда название Pelycosauria[33][34]. С другой стороны, объём Anomodontia постепенно был ограничен лишь дицинодонтами и близкородственными группами[35].
XX век и начало XXI века
На протяжении XX века и в начале XXI века изучение синапсид продолжалось. Были описаны новые роды и виды, а на основании анализа особенностей их скелета, микроструктуры костей и зубной системы получены новые сведения о морфологии, физиологии и образе жизни синапсид[36][37]. Стало практически общепринятым деление пеликозавров на 6 семейств (Ophiacodontidae, Varanopidae, Eothyrididae, Caseidae, Edaphosauridae, Sphenacodontidae)[38][39] и терапсид на 6 основных групп (Biarmosuchia, Dinocephalia, Anomodontia, Gorgonopsia, Therocephalia, Cynodontia)[35], причём в основном утвердился консенсус относительно филогенетических связей между перечисленными группами синапсид (см. раздел «Классификация синапсид»). Особое место в системе синапсид заняли раннепермские роды тетрацератопс (Tetraceratops, обнаружен в 1897 и описан в 1908 году) и раранимус (Raranimus, обнаружен в 1998 и описан в 2009 году), которые совмещают в себе признаки, характерные для пеликозавров и терапсид; эти два рода, видимо, образуют соответственно две наиболее ранние (последовательно отделившиеся) ветви терапсид[35][40][41].

В конце XX века получила распространение трактовка таксона Synapsida как самостоятельного класса (а не подкласса в составе класса Reptilia, как раньше)[7][8]; заметим, что Адриан Бринк ещё в 1963 году предлагал[42] поднять таксономический ранг Synapsida до класса с двумя подклассами: Pelycosauria и Therapsida[43]. В то же время распространение кладистики привело к тому, что теперь название Synapsida обычно используется в расширенном смысле и применяют его к кладе, включающей не только синапсид в классическом понимании, но и млекопитающих (по отношению к данной кладе реже применяют также названия Theropsida[10][44][45] или Pan-Mammalia[46]). При этом млекопитающих также вводят и в состав Therapsida, а таксон Pelycosauria не используют — как относящийся к парафилетической группе[35][47].
Особенности строения
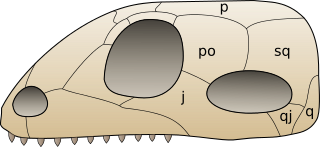
Характерной чертой синапсид является наличие с каждой стороны их черепа одного височного отверстия, ограниченного сверху височной дугой из заглазничной и чешуйчатой костей (у завроптеригий — тоже одно височное отверстие, но лежащее выше заглазничной кости, а у диапсид — два височных отверстия). На приведённом справа рисунке показано типичное для синапсид строение черепа и обозначены кости: p — теменная, po — заглазничная, sq — чешуйчатая, j — скуловая, qj — квадратно-скуловая, q — квадратная[48]. Височные отверстия служат для выхода челюстных мышц, а их наличие позволяло тем амниотам, которые их имели, более эффективно — по сравнению с земноводными и черепахами (череп которых височных отверстий не имеет) — кусать свою добычу.
Нижняя челюсть у большинства немаммальных синапсид имеет полный набор костей, характерный для ранних амниот. С черепом она сочленяется челюстным суставом, образованным (как и у других амниот, но не у млекопитающих) квадратной и сочленовной костями. У млекопитающих же на смену такому сочленению приходит вторичный челюстной сустав, образуемый уже чешуйчатой и зубной костями[49][50]. Традиционно именно формирование вторичного челюстного сустава рассматривают как ведущий признак, позволяющий провести грань между классами млекопитающих и синапсид; уточнённая формулировка диагностического признака млекопитающих, предложенная в 2002 году Ло Чжэси, З. Келян-Яворовской и Р. Чифелли, звучит так: «сочленение нижней челюсти с черепом, при котором сочленовная головка зубной кости входит в гленоид (суставную впадину) на чешуйчатой кости» (у синапсид из семейства Tritheledontidae тоже сочленяются чешуйчатая и зубная кости, хотя нет ни сочленовной головки, ни выраженной области гленоида)[51].
Уже у некоторых ранних синапсид (пеликозавров) зубы обнаруживают начальные этапы дифференцировки: зубы, располагавшиеся на предчелюстных костях, напоминают по форме резцы, далее следуют клыки и щёчные зубы (последние сохраняют исходную коническую форму)[52]. Более выраженный характер дифференцировка зубов приобретает у продвинутых синапсид (терапсид); чётко выражена она у представителей группы териодонтов («зверозубые»), причём если у горгонопсов щёчные зубы остаются простыми коническими (а иногда вовсе исчезают), то у цинодонтов они имеют сложно устроенную жевательную поверхность[53].
Важным эволюционным завоеванием продвинутых териодонтов стало формирование вторичного костного нёба (оно происходило независимо у триасовых тероцефалов и цинодонтов). Вторичное нёбо существенно укрепило верхнюю челюсть и надёжно обособило носовую и ротовую полости, позволяя животному непрерывно дышать при пережёвывании пищи[54][55].
Судя по всему, синапсиды, в отличие от завропсид, изначально имели гладкую, лишённую чешуи кожу, в которой имелись многочисленные железы. Для большинства млекопитающих и, вероятно, некоторых их вымерших родственников, характерен волосяной покров, обеспечивавший термоизоляцию (полагают, что волосы первоначально могли появиться в виде средств осязания — таких, как вибриссы[56]). Немаммальные синапсиды размножались путём откладывания яиц[57]. Среди млекопитающих однопроходные также сохранили эту особенность; все остальные современные млекопитающие являются живородящими[58].
Классификация синапсид
Традиционно в состав класса синапсид включают два отряда — Pelycosauria (пеликозавры, средний карбон — поздняя пермь) и Therapsida (терапсиды, средний пермь — ранний мел); последний из них включает такие основные группы: Biarmosuchia (биармозухии), Dinocephalia (диноцефалы), Anomodontia (аномодонты) и Theriodontia (териодонты, или зверозубые)[59][60]. Некоторые авторы повышают ранг терапсид до подкласса, содержащего отряды Biarmosuchia, Dinocephalia, Anomodontia и надотряд Theriodontia; последний при этом, в свою очередь, включает отряды Gorgonopsia (горгонопсы) и Eutheriodontia (настоящие териодонты)[61]. Согласно современным данным[39][62], классификация синапсид может быть представлена в виде следующих двух кладограмм:
| Synapsida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Therapsida |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Из этих кладограмм видно, что оба отряда в традиционном объёме (как и весь класс Synapsida) являются парафилетическими группами.
Для детального прослеживания происхождения млекопитающих приведём ещё одну кладограмму, раскрывающую внутреннюю структуру клады Cynodontia (цинодонты) и составленную с учётом результатов исследования Лю Цзюня и П. Олсена (2010)[65]:
| Cynodontia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционная история

Отделение синапсидной ветви амниот от завропсидной их ветви произошло, по современным данным, 310 млн лет тому назад (средний карбон) или несколько раньше; верхняя граница для времени отделения оценивается при этом как 333 млн лет назад (нижний карбон)[66][67].
В качестве одного из древнейших представителей синапсид обычно рассматривают археотириса (Archaeothyris) из семейства Ophiacodontidae, жившего 306 млн лет назад (средний карбон) и описанного в 1972 году по ископаемым остаткам, найденных в Новой Шотландии. К синапсидам относят также раннюю амниоту Protoclepsydrops, жившую несколько раньше (314 млн лет назад) и описанную в 1964 году по фрагментарным остаткам (также из Новой Шотландии); его также (предположительно) относят к Ophiacodontidae, однако систематическое положение данного рода остаётся неясным[45][68]. Внешне ранние синапсиды напоминали крупных современных ящериц типа игуан или варанов, но с относительно более короткими конечностями (примером может служить варанозавр — Varanosaurus, живший в конце ранней перми активный хищник средних размеров из того же семейства Ophiacodontidae[69])[59].

К началу перми синапсиды составляли до 70 % известных родов амниот; к этому времени они уже дивергировали на ряд самостоятельных семейств (включавших рыбоядных, растительноядных и плотоядных животных[39]). При этом в ранней перми представители семейства Sphenacodontidae заняли доминирующие позиции в качестве наземных хищников. Наибольшую известность среди них приобрёл диметродон (Dimetrodon), на спине которого располагался «парус» из кожной перепонки и поддерживавших её многочисленных остистых отростков, который, скорее всего, служил целям терморегуляции (аналогичным «парусом» обладали и некоторые Edaphosauridae; представители данного семейства заняли трофическую нишу крупных специализированных фитофагов)[70][71]. По расчётам К. Брамуэлла и П. Феллгетта, 200-килограммовый диметродон разогревался бы без паруса с 26 °C до 32 °C за 205 минут, а с парусом — за 80 минут[72].
На рубеже ранней и средней перми на смену пеликозаврам приходят терапсиды, отличавшиеся существенно более активным образом жизни и высоким уровнем метаболизма и занимавшие в течение средней и поздней перми доминирующие позиции среди наземных позвоночных[39][73]. Они быстро вытеснили пеликозавров, и лишь немногие из последних дожили до конца пермского периода. Некоторое исключение составляют казеиды (Caseidae), переживавшие расцвет именно в средней перми и занявшие ту трофическую нишу крупных специализированных фитофагов, которую в начале периода занимали эдафозавры[39][71].
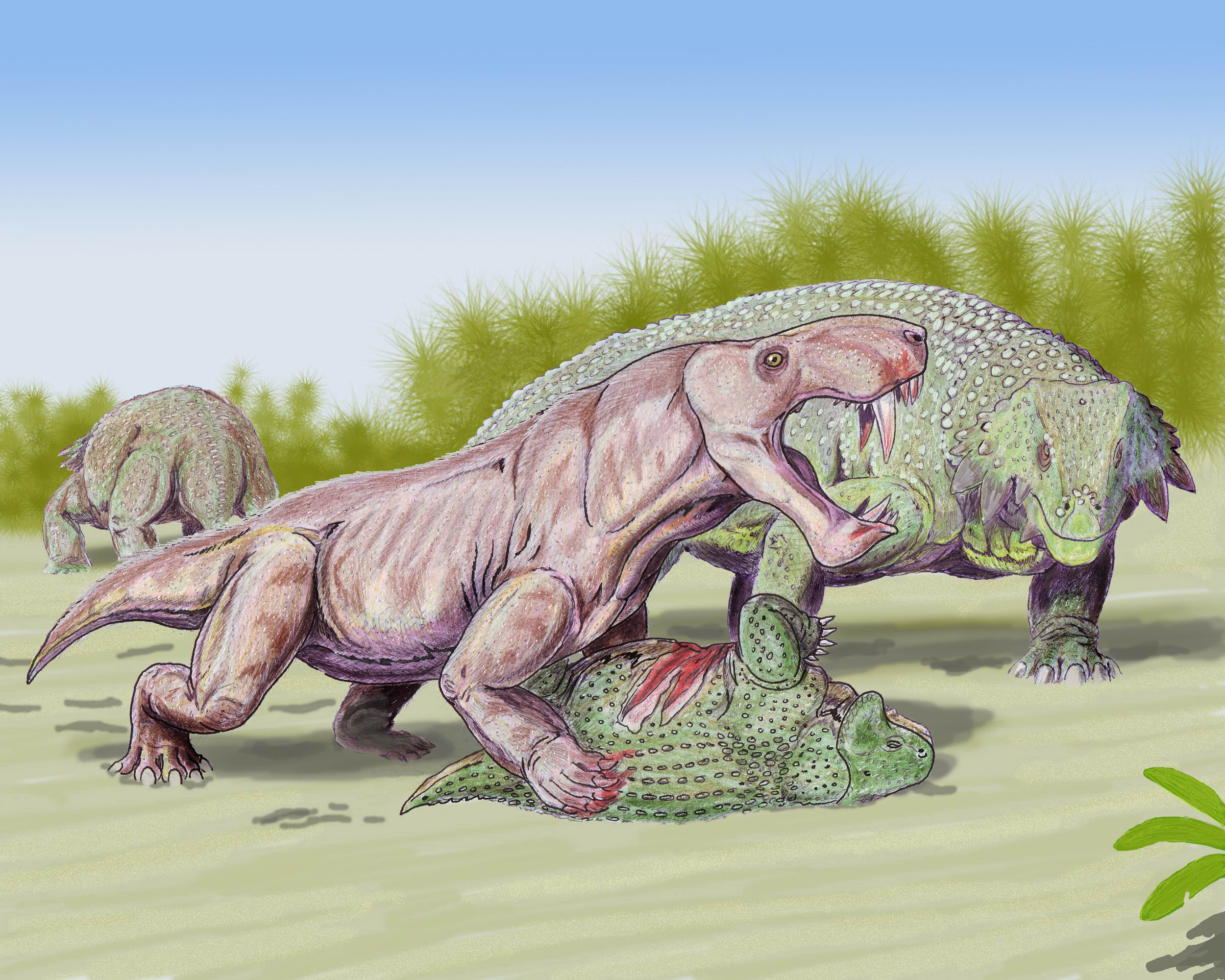
Терапсиды достаточно быстро дивергировали на ряд групп; среди них хищный образ жизни вели биармозухии (Biarmosuchia), тероцефалы (Therocephalia) и ранние цинодонты (Cynodontia), а дейноцефалы (Dinocephalia) включали как хищные, так и растительноядные формы[74]. Фитофагами были и дицинодонты (Dicynodontia), разнообразные по размерам и строению тела. Роль доминирующих наземных хищников теперь играли горгонопсы (Gorgonopsia), охотившиеся на крупную добычу; они вцеплялись в неё большими саблевидными клыками, а затем разрывали жертву резкими продольными рывками[75]. Одним из наиболее крупных горгонопсов была иностранцевия (Inostrancevia), длина черепа которой могла достигать 60 см[76][77].
В конце пермского периода (около 252 млн лет назад[78]) произошло катастрофическое пермское вымирание, в результате которого исчезло примерно 90 % видов морских животных и около 70 % семейств наземных животных. Вымирание затронуло и синапсид: исчезли последние пеликозавры, а среди синапсид катастрофу пережили лишь представители трёх групп — крупные растительноядные дицинодонты и мелкие (вероятно, насекомоядные или всеядные) тероцефалы и цинодонты[79].

Tritylodon longaevus
В триасе тероцефалы, впрочем, были относительно немногочисленными и исчезли к концу средней части периода[80]. Более широким распространением на протяжении триаса отличались дицинодонты, хотя значительную диверсификацию в течение периода претерпела лишь одна (клада Kannemeyeriiformes) из четырёх их групп, переживших пермское вымирание; к концу триаса исчезают и дицинодонты (хотя, по некоторым данным, на территории Австралии они просуществовали вплоть до конца раннего мела[81])[82]. Зато подлинный расцвет в триасе переживают цинодонты, и именно к этому периоду относится большинство их семейств. Большинство цинодонтов оставались хищными или насекомоядными, однако появились и специализированные растительноядные группы: гомфодонты (Gomphodontia) и тритилодонты (Tritylodontydae). У триасовых цинодонтов возникает значительное число признаков, характерных для млекопитающих[83].
В позднем триасе появляются первые млекопитающие. Большинство эволюционных линий немаммальных цинодонтов прекращают существование к концу триаса. В юрском периоде встречались представители немногих семейств: семейства Traversodontidae из группы гомфодонтов (представлено в ранней юре родом Scalenodontoides) и филогенетическими близких к млекопитающим семейств Tritheledontidae (закончило существование в средней юре) и Tritylodontydae (его представители дожили до конца раннего мела, где представлены родом Xenocretosuchus[84])[85][86].
Примечания
Комментарии
Источники
- Сина́псиды // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов. — М. : Большая российская энциклопедия, 2004—2017.
- Еськов К. Ю. Словарь названий живых и ископаемых организмов // Удивительная палеонтология: история Земли и жизни на ней. — М.: ЭНАС, 2008. — С. 303. — 312 с. — ISBN 978-5-91921-129-7.
- Проверка слова «синапсиды». Gramota.ru, справочно-информационный интернет-портал «Русский язык». Дата обращения: 13 июня 2020. Архивировано 7 июля 2018 года.
- Laurin, Michel, and Robert R. Reisz. Synapsida: Mammals and their extinct relatives. The Tree of Life Web Project (2011). Архивировано 7 декабря 2012 года.
- Romer A. S. & Parsons T. S. (1985). The Vertebrate Body. (6th ed.) Saunders, Philadelphia.
- Кэрролл, т. 3, 1993, с. 203—205.
- Benton, 2005, p. 394.
- Vaughan, Ryan, Czaplewski, 2011, p. 47.
- White A. T. Reptilomorpha: Cotylosauria (section Amniota). // Website «PALAEOS: The history of life on Earth». Дата обращения: 3 мая 2016. Архивировано 20 декабря 2010 года.
- Rieppel O. Monophyly, Paraphyly, and Natural Kinds (англ.) // Biology and Philosophy. — 2005. — Vol. 20 , no. 2. — P. 465—487. — doi:10.1007/s10539-004-0679-z. Архивировано 3 декабря 2018 года..
- Synapsida (англ.) информация на сайте «Энциклопедия жизни» (EOL) (Дата обращения: 3 октября 2020).
- Synapsida (англ.). The Taxonomicon. Дата обращения: 2 октября 2020. Архивировано 30 октября 2020 года.
- Palmer, 2005, p. 153.
- Ochev, Surkov, 2003, p. 1.
- Ивахненко, 2008, с. 101, 118—125.
- Palmer, 2005, p. 154.
- Ochev, Surkov, 2003, p. 2.
- Fröbisch J. . Anomodontia: Introduction // Early Evolutionary History of the Synapsida / Ed. by C. F. Kammerer, K. D. Angielczyk, J. Fröbisch. — Dordrecht: Springer Science & Business Media, 2013. — xi + 337 p. — (Vertebrate Paleobiology and Paleoanthropology Series). — ISBN 978-94-007-6841-3. — P. 89—92.
- Brink K. S., Maddin H. C., Evans D. C., Reisz R. R. . Re-evaluation of the historic Canadian fossil Bathygnathus borealis from the Early Permian of Prince Edward Island // Canadian Journal of Earth Sciences, 2015, 52 (12). — P. 1109—1120. — doi:10.1139/cjes-2015-0100.
- Reisz R. R. . «Pelycosaur»-Grade Synapsids: Introduction // Early Evolutionary History of the Synapsida / Ed. by C. F. Kammerer, K. D. Angielczyk, J. Fröbisch. — Dordrecht: Springer Science & Business Media, 2013. — xi + 337 p. — (Vertebrate Paleobiology and Paleoanthropology Series). — ISBN 978-94-007-6841-3. — P. 3—5.
- Ивахненко, 2008, с. 101.
- Zittel K. A. . History of Geology and Palæontology to the End of the Nineteenth Century. — London: Walter Scott, 1901. — xiii + 561 p. Архивная копия от 5 июня 2016 на Wayback Machine — P. 517.
- Hay O. P. . Bibliography and Catalogue of the Fossil Vertebrata of North America. — Washington: U. S. Government Printing Office, 1902. — 868 p. — (Bulletin of the United States Geological Survey, no. 179). Архивная копия от 5 июня 2016 на Wayback Machine — P. 426.
- Hu Yaoming, Meng Jin, Clark J. M. . A new tritylodontid from the Upper Jurassic of Xinjiang, China // Acta Palaeontologica Polonica, 2009, 54 (12). — P. 385—391. — doi:10.4202/app.2008.0053.
- Kemp T. S. . The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis // Journal of Evolutionary Biology, 2006, 19 (4). — P. 1231—1247. — doi:10.1111/j.1420-9101.2005.01076.x.
- Gow C. E. . The importance of Karoo fossils in the search for mammal origins // Palaeontologia africana, 1997, 33. — P. 23—27.
- Кэрролл, т. 2, 1993, с. 188, 194—197, 200—201, 208—213, 219.
- Baur G., Case E. C. . The History of the Pelycosauria, with a Description of the Genus Dimetrodon, Cope // Transactions of the American Philosophical Society. New Series, 1889, 20 (1). — P. 5—62. — doi:10.2307/1005488.
- Ивахненко М. Ф. . Тетраподы Восточно-Европейского плакката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001. — 200 с. — (Труды Палеонтологического института РАН, т. 283). — ISBN 5-88345-064-4. Архивная копия от 5 июня 2016 на Wayback Machine — С. 19—20.
- Ochev, Surkov, 2003, p. 2—5.
- Osborn H. F. . The Reptilian Subclasses Diapsida and Synapsida and the Early History of the Diaptosauria // Memoirs of the AMNH, 1903, 1 (8). — P. 1109—1120.
- Romer A. S., Price L. I. . Review of the Pelycosauria. — New York: Geological Society of America, 1940. — x + 538 p. — (Geological Society of America. Special papers, no. 28).
- Williston S. W. . The Osteology of the Reptiles. — Cambridge: Harvard University Press, 1925. — xiii + 300 p. — P. 211, 228, 236—237.
- Spindler F., Scott D., Reisz R. R. . New Information on the Cranial and Postcranial Anatomy of the Early Synapsid Ianthodon schultzei (Sphenacomorpha: Sphenacodontia), and its Evolutionary Significance // Fossil Record, 2015, 18. — P. 17—30. — doi:10.5194/fr-18-17-2015.
- Rubidge B. S., Sidor C. A. . Evolutionary Patterns among Permo-Triassic Therapsids // Annual Review of Ecology and Systematics, 2001, 32. — P. 449—480. — doi:10.1146/annurev.ecolsys.32.081501.114113.
- Кэрролл, т. 2, 1993, с. 222—224.
- Huttenlocker A. K., Rega E. . The Paleobiology and Bone Microstructure of Pelycosaurian-Grade Synapsids // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 91—120.
- Benton, 2005, p. 120—124.
- Kemp T. S. . The Origin and Radiation of Therapsids // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 3—30.
- Amson E., Laurin M. . On the Affinities of Tetraceratops insignis, an Early Permian Synapsid // Acta Palaeontologica Polonica, 2011, 56 (2). — P. 301—312. — doi:10.4202/app.2010.0063.
- Liu Jun, Rubidge B. S., Li Jinling. . New Basal Synapsid Supports Laurasian Origin for Therapsids // Acta Palaeontologica Polonica, 2009, 54 (3). — P. 393—400. — doi:10.4202/app.2008.0071.
- Brink A. S. . The taxonomic position of the Synapsida // South African Journal of Science, 1963, 59 (5). — P. 153—159.
- Ивахненко, 2008, с. 102.
- Marjanović D., Laurin M. . Fossils, Molecules, Divergence Times, and the Origin of Lissamphibians // Systematic Biology, 2007, 56 (3). — P. 369—388. — doi:10.1080/10635150701397635.
- Kermack D. M., Kermack K. A. . The Evolution of Mammalian Characters. — Dordrecht: Springer Science & Business Media, 2013. — x + 149 p. — ISBN 978-1-4214-0040-2. Архивная копия от 5 июня 2016 на Wayback Machine — P. 10—13.
- Sudhaus W. . From the Cladogram to an Explanation of Anagenesis in an Evolutionary History Perspective, Exemplified by the Mammals // Peckiana, 2016, 11. — P. 43—65. — doi:10.1080/10635150701397635.
- Linzey D. W. . Vertebrate Biology. 2nd ed. — Baltimore: The Johns Hopkins University Press, 2012. — xiv + 583 p. — ISBN 978-1-4214-0040-2. Архивная копия от 5 июня 2016 на Wayback Machine — P. 269—271.
- Черепанов, Иванов, 2007, с. 186, 251.
- Черепанов, Иванов, 2007, с. 251.
- Дзержинский, Васильев, Малахов, 2014, с. 435, 438.
- Luo Zhe-Xi, Kielan-Jaworowska, Cifelli, 2002, p. 19—20.
- Дзержинский, Васильев, Малахов, 2014, с. 435.
- Черепанов, Иванов, 2007, с. 255, 259—261.
- Дзержинский, Васильев, Малахов, 2014, с. 369, 438—439.
- Кэрролл, т. 2, 1993, с. 196—197.
- Дзержинский, Васильев, Малахов, 2014, с. 439.
- Черепанов Г. О., Иванов А. О. Ископаемые высшие позвоночные. 2-е изд. — СПб.: Изд-во С.-Петербургского ун-та, 2007. — 202 с. — ISBN 978-5-288-04308-6. — С. 103.
- Gerardo De Iuliis, Dino Pulerà. The Dissection of Vertebrates (англ.). — 3nd ed. — Academic Press, 2019. — P. 21. — 394 p. — ISBN 978-0-12-410460-0.
- Кэрролл, т. 2, 1993, с. 176.
- Черепанов, Иванов, 2007, с. 255.
- Черепанов, Иванов, 2007, с. 255, 259.
- Laurin M., Reisz R. R. Synapsida. Mammals and their extinct relatives Архивная копия от 7 декабря 2012 на Wayback Machine. The Tree of Life Web Project (2011).
- Ford D. P., Benson R. B. J. A redescription of Orovenator mayorum (Sauropsida, Diapsida) using high-resolution μCT, and the consequences for early amniote phylogeny (англ.) // Papers in Palaeontology : journal. — 2019. — Vol. 5, iss. 2. — P. 197—239. — ISSN 2056-2802. — doi:10.1002/spp2.1236.
- Ford D. P., Benson R. B. J. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae (англ.) // Nature Ecology & Evolution : journal. — 2020. — Vol. 4, iss. 1. — P. 57—65. — ISSN 2397-334X. — doi:10.1038/s41559-019-1047-3.
- Liu Jun, Olsen P. The Phylogenetic Relationships of Eucynodontia (Amniota: Synapsida) // Journal of Mammalian Evolution, 2010, 17 (3). — P. 151—176. — doi:10.1007/s10914-010-9136-8.
- Oftedal O. T. . The Mammary Gland and Its Origin during Synapsid Evolution // Journal of Mammary Gland Biology and Neoplasia, 2002, 7 (3). — P. 225—252. — doi:10.1023/A:1022896515287. — PMID 12751889.
- Benton M. J., Donoghue P. C. J., Asher R. J., Friedman M., Near T. J., Vinther J. . Constraints on the Timescale of Animal Evolutionary History // Palaeontologia Electronica, 2015, 18.1.1FC. — P. 1—106.
- Benton M. J., Donoghue P. C. J. . Paleontological Evidence to Date the Tree of Life. — Molecular Biology and Evolution, 2007, 24 (1). — P. 26—53. — doi:10.1093/molbev/msl150.
- Cloudsley-Thompson J. L. . Ecology and Behaviour of Mesozoic Reptiles. — Heidelberg: Springer Science & Business Media, 2005. — xii + 219 p. — (Lecture Notes in Statistics, vol. 181). — ISBN 3-540-22421-1. Архивная копия от 24 апреля 2016 на Wayback Machine — P. 14.
- Кэрролл, т. 2, 1993, с. 178, 180.
- Черепанов, Иванов, 2007, с. 254.
- Bramwell C. D., Fellgett P. P. . Thermal Regulation in Sail Lizards. — Nature, 1973, 242 (5395). — P. 203—205. — doi:10.1038/242203a0.
- Дзержинский, Васильев, Малахов, 2014, с. 435—436.
- Benton, 2005, p. 125, 130, 292.
- Дзержинский, Васильев, Малахов, 2014, с. 437.
- Константинов В. М., Наумов С. П., Шаталова С. П. . Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5. — С. 192.
- Черепанов, Иванов, 2007, с. 259.
- Shen Shuzhong, Crowley J. L., Wang Yue, Bowring S. A., Erwin D. H., Sadler P. M., Cao Changqun, Rothman D. H., Henderson C. M., Ramezani J., Zhang Hua, Shen Yanan, Wang Xiangdong, Wang Wei, Mu Lin, Li Wenzhong, Tang Yuegang, Liu Xiaolei, Liu Lujun, Zeng Yong, Jiang Yaofa, Jin Yugan. . Calibrating the End-Permian Mass Extinction. — Science, 2011, 334 (6061). — P. 1367—1372. — doi:10.1126/science.1213454. — PMID 22096103.
- Smith R., Rubidge B., van der Valt M. . Therapsid Bioiversity Patterns and Paleoenvironments of the Karoo Basin, South Africa // Forerunners of Mammals: Radiation, Histology, Biology / Ed. by A. Chinsamy-Turan. — Bloomington, Indiana: Indiana University Press, 2011. — xv + 330 p. — ISBN 978-0-253-35697-0. — P. 31—64.
- Abdala F., Rubidge B. S., Van Den Heever J. . The Oldest Therocephalians (Therapsida, Eutheriodontia) and the Early Diversification of Therapsida. — Palaeontology, 2008, 51 (4). — P. 1011—1024. Архивная копия от 4 мая 2016 на Wayback Machine — doi:10.1111/j.1475-4983.2008.00784.x.
- Thulborn T., Turner S. . The Last Dicynodont: An Australian Cretaceous Relict. — Proc. of the Royal Society: Biological Sciences, 2003, 270 (1518). — P. 985—993. Архивная копия от 19 апреля 2016 на Wayback Machine — doi:10.1098/rspb.2002.2296.
- Kammerer C. F., Fröbisch J., Angielczyk K. D. . On the Validity and Phylogenetic Position of Eubrachiosaurus browni, a Kannemeyeriiform Dicynodont (Anomodontia) from Triassic North America. — PLoS One, 2013, 8 (5): e64203. Архивная копия от 12 октября 2017 на Wayback Machine — doi:10.1371/journal.pone.0064203.
- Benton, 2005, p. 289—292.
- Лопатин А. В, Агаджанян А. К. Тритилодонт (Tritylodontidae, Synapsida) из мезозоя Якутии // Доклады Академии наук. — 2008. — Т. 419, № 2. — С. 279—282. Архивировано 11 октября 2017 года.
- Benton, 2005, p. 297—298.
- Ruta M., Botha-Brink J., Mitchell S. A., Benton M. J. . The Radiation of Cynodonts and the Ground Plan of Mammalian Morphological Diversity // Proc. of the Royal Society B, 2013, 280 (1769), 20131865. — doi:10.1098/rspb.2013.1865.
Литература
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. . Зоология позвоночных. — 2-е изд. — М.: Издат. центр «Академия», 2014. — 464 с. — ISBN 978-5-4468-0459-7.
- Ивахненко М. Ф. . Подкласс Theromorpha // Ископаемые рептилии и птицы. Ч. 1 / отв. ред. М. Ф. Ивахненко, Е. Н. Курочкин. — М.: ГЕОС, 2008. — С. 101—183. — 348 с. — (Ископаемые позвоночные России и сопредельных стран). — ISBN 978-5-89118-415-6.
- Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3-х тт. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.
- Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3-х тт. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0.
- Черепанов Г. О., Иванов А. О. Палеозоология позвоночных. — М.: Издат. центр «Академия», 2007. — 352 с. — ISBN 978-5-7695-3104-0.
- Benton M. J. Vertebrate Palaeontology. 3rd ed. — Oxford: Blackwell Science Ltd, 2005. — 455 p. — ISBN 0-632-05637-1.
- Colbert E. H. . Evolution of the Vertebrates. 2nd ed. — N. Y.: John Wiley & Sons, 1969. — 477 p.
- Luo Zhe-Xi, Kielan-Jaworowska Z., Cifelli R. L. . In Quest for a Phylogeny of Mesozoic Mammals. — Acta Palaeontologica Polonica, 2002, 47 (1). — P. 1—78.
- Ochev V. G., Surkov V. V. . The history of excavation of Permo-Triassic vertebrates from Eastern Europe // The Age of Dinosaurs in Russia and Mongolia / Ed. by M. J. Benton, M. A. Shishkin, D. M. Unwin, E. N. Kurochkin. — Cambridge: Cambridge University Press, 2003. — xxxv + 740 p. — ISBN 0-521-55476-4. — P. 1—16.
- Palmer D. . Earth Time: Exploring the Deep Past from Victorian England to the Grand Canyon. — Chichester: John Wiley & Sons, 2005. — xix + 436 p. — ISBN 0-470-02221-3.
- Vaughan T. A., Ryan J. M., Czaplewski N. J. . Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
Ссылки
- Laurin M., Reisz R. R. Tree of Life — Synapsida. 1997.
- Synapsida — Pelycosauria (англ.)
- Transitional Vertebrate Fossils (англ.)
- Синапсиды — Вымершие животные вики (рус.)
| Таксономия | |
|---|---|
| В библиографических каталогах |
|
| Классы хордовых (Chordata) | |
|---|---|
| |
| Бесчерепные (Acrania) | |
| Olfactores | |
† — вымерший таксон, * — парафилетический таксон | |
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||




На других языках
[es] Synapsida
Los sinápsidos (Synapsida, griego "arco fusionado") también denominados Theropsidos, son una clase de amniotas que incluye a los mamíferos y a todas aquellas formas más relacionadas con ellos que con el resto de amniotas.[1] Los sinápsidos no mamíferos se han denominado tradicionalmente "reptiles mamiferoides", es decir reptiles similares a los mamíferos.[2][3]- [ru] Синапсиды
Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии