bio.wikisort.org - Animal
Euchambersia est un genre éteint de thérapsides thérocéphales ayant vécu durant la fin du Permien (Wuchiapingien), il y a entre 256 et 255 millions d'années avant notre ère, dans ce qui sont actuellement l'Afrique du Sud et la Chine. L'espèce type, Euchambersia mirabilis, est nommé par le paléontologue Robert Broom en à partir d'un crâne auquel manque la mâchoire inférieure. Un deuxième crâne attribué à l'espèce, appartenant probablement à un individu juvénile et toujours dans le même état de conservation, a été décrit par la suite. En , une deuxième espèce, Euchambersia liuyudongi, est officiellement décrite par les paléontologues Jun Liu et Fernando Abdala à partir d'un crâne bien préservé et de quelques restes postcrâniens fragmentaires. Le genre appartient à la famille éteinte des Akidnognathidae, historiquement désignée sous le synonyme Euchambersiidae (nommé d'après Euchambersia).

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Therocephalia |
| Famille | † Akidnognathidae |
Genre
Broom, 1931
Espèces de rang inférieur
- † E. mirabilis (type) Broom, 1931
- † E. liuyudongi Liu & Abdala, 2022
Euchambersia est un thérocéphale de petite taille disposant d'un museau court et de grandes canines typique du groupe. Cependant, il se distingue parmi les thérocéphales de par la présence de rainures sur les canines et d'une grande indentation sur le côté du crâne. Certains auteurs considèrent que la rainure peu profonde situé à l'arrière des canines ferait partie d'un mécanisme de distribution de venin, mais cette hypothèse est mal étayée. Cependant, si cette supposition s'avère vraie, alors il s'agirait donc d'un des plus anciens tétrapodes connus se à disposer de telles caractéristiques. Une étude publié en 2017 utilise la structure interne du crâne d'E. mirabilis comme preuve la plus forte en faveur de l'hypothèse concernant la venimosité. D'autres possibilités, comme l'existence d'une sorte d'organe sensoriel, restent encore plausibles pour expliquer la présence de certaines structures.
Découvertes et étymologie
L'holotype du genre Euchambersia, plus précisément de l'espèce type Euchambersia mirabilis, est découvert par le paléontologue Robert Broom durant le début du XXe siècle dans la ferme de Vanwyksfontein, propriété d'un certain M. Greathead, près de la ville de Norvalspont (en), au Cap-Nord, en Afrique du Sud. Il se compose d'un seul crâne déformé, catalogué NHMUK R5696, décrit plus en détail par Broom dans un article qu'il publie en [1]. C'est en 1966 qu'un deuxième crâne attribué à la même espèce, catalogué BP/1/4009, plus petit, y est trouvé, avant d'être décrit officiellement par James Kitching onze ans après, en [2],[3],[4]. Il manque la mâchoire inférieure aux deux spécimens. Ceux-ci proviennent de la même couche générale de roche, dans la zone d'assemblage supérieure de Cistecephalus du groupe de Beaufort, au sein du supergroupe du Karoo[3]. Ce site est daté du stade Wuchiapingien du Permien supérieur[5], entre 256,2 et 255,2 millions d'années[6].
Le genre Euchambersia, qualifié par Robert Broom comme « le thérocéphale le plus remarquable jamais découvert », est nommé par ce dernier d'après l'éminent éditeur écossais et penseur évolutionniste Robert Chambers, dont Broom considère le livre Vestiges of the Natural History of Creation comme « un travail remarquable [bien que] méprisé par beaucoup »[1]. L'épithète spécifique du taxon, venant du latin mirabilis, signifiant « merveilleux », est nommé en raison de l'aspect unique de l'animal qui le différencie des autres thérocéphales[1].
La deuxième espèce, Euchambersia liuyudongi, est nommée par les paléontologues Jun Liu et Fernando Abdala en sur la base d'un crâne bien conservé mesurant 7 centimètres de long. Contrairement aux deux spécimens connus d'E. mirabilis, le spécimen catalogué IVPP V 31137 se dispose d'une mâchoire inférieure associée et de quelque restes postcrâniens qui inclut six vertèbres et quelques fragments de côtes. Il est originaire de la formation de Naobaogou, en Mongolie intérieure (Chine), un site datant de la même époque à celui d'où est connu l'espèce sud-africain. L'épithète spécifique est nommé en l'honneur d'un technicien ayant découvert le spécimen[7].
Description
Description simplifié

Euchambersia est un thérapside de petite taille possédant un museau relativement court pour un thérocéphale, le crâne holotype, conséquence de la déformation et de l'écrasement lors de la fossilisation, ayant une longueur déduite d'environ 11,6 cm. Le deuxième crâne connu, d'une longueur de 8,2 cm[2],[3], appartient à un individu plus petit, très probablement à un individu juvénile, à en juger par le manque de fusion dans le crâne[4]. Le crâne de la deuxième espèce mesure 7 centimètres de longueur, mais il n'est pas précisé si le spécimen appartient à un adulte ou à un individu juvénile[7].
Selon les premières descriptions, les orbites des crânes d'Euchambersia sont plutôt petites. Les branches des os postorbitaires et de l'os jugal, qui entourent habituellement l'arrière et le bas de l'orbite oculaire chez les thérocéphales, semblent être soit très réduites, soit totalement absentes. Le haut de l'orbite est formé par l'os préfrontal et le frontal est également petit. Le crâne ne porte pas de foramen pinéal. Comme Theriognathus, le ptérygoïde et le palatin du palais ne sont pas séparés, par aucune sorte d'ouverture, du transpalatin, situé plus loin sur le côté de la mâchoire[1].
Dentition
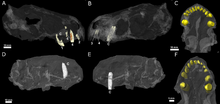
Bien que les crânes soient incomplètement préservés, la tomodensitométrie suggère que chaque prémaxillaire contient cinq incisives, les alvéoles devenant progressivement plus grandes de la première à la cinquième. Comme chez les autres thériodontes, les couronnes des incisives d'Euchambersia sont coniques et manque également de dentelures, contrairement aux gorgonopsiens et à certains autres thérocéphales scylacosauriens, groupe dont le genre en fait partie. Le bord intérieur des incisives paraît être légèrement concave et le bord arrière semble avoir une crête. Le second spécimen a une incisive déplacée, conservée dans la cavité nasale, plus fortement recourbée et présentant des marques d'usure sur le bord supérieur, suggérant qu'il s'agit d'une incisive provenant de la mâchoire inférieure. La quatrième incisive a également une dent de remplacement qui pousse derrière elle, accompagnée d'une résorption (en) de la racine[4].
Le spécimen type préserve la canine droite[4]. Comme chez d'autres thérocéphales, les canines d'Euchambersia sont proportionnellement très grandes, ce qui suggère un mode de vie de prédateur spécialisé qui infligerait une morsure similaire à celle d'un animal à « dents de sabre » pour l'abattage des proies[8]. Cette canine est ronde en coupe transversale, et porte une crête proéminente sur le côté de la surface avant[3]. Immédiatement à côté de cette crête se trouve une dépression peu profonde qui s'élargit près du sommet de la dent, qui est probablement la même structure que la rainure interprétée par certains auteurs[4],[9]. Les thériodontes remplacent généralement leurs dents en alternance de telle sorte que la canine soit toujours fonctionnelle[10],[11],[12]. Les deux crânes connus d'E. mirabilis ne montrent aucun signe de développement de canines de remplacement, ce qui suggère que l'animal dépendait de la présence et de la fonctionnalité simultanée de ces dernières[4].
Fosse maxillaire et canaux associés

Aucune dent supplémentaire n'est présente derrière les incisives et les canines supérieures. Là où les thérocéphales ont des dents derrière les canines, se trouve à la place une grande dépression, ou fosse, du côté du maxillaire ; celle-ci est également délimitée en dessous par une partie du lacrymal et peut-être une partie du jugal[1]. Chez, E. mirabilis, cette fosse mesure 48 % de la longueur de la mâchoire dans le spécimen type et 38 % dans le second crâne. Dans ces crânes, cette fosse est divisée en deux parties : une crête moins profonde sur le dessus et une dépression plus grande et plus profonde sur le fond. Un large sillon commençant derrière la canine entre en contact avec le fond de la fosse et passe ensuite à l'intérieur de la bouche. La partie inférieure de la fosse est fortement piquée et porte une petite ouverture, ou foramen, sur les surfaces avant et arrière[4].
La tomodensitométrie montre que ces ouvertures conduisent à des canaux. Ceux-ci se connectent au nerf trijumeau, qui contrôle la sensibilité faciale. Le canal dirigé vers l'avant se divise également en trois branches principales du nerf infra-orbitaire, qui se connectent toutes à l'alvéole de la canine, la jonction se produisant à environ 3 à 6 mm le long du canal, un autre point de variation entre les deux crânes connus d'E. mirabilis[13]. La branche supérieure, la branche nasale externe, se divise en quatre branches dans le crâne holotype, mais elle ne se divise pas dans le deuxième crâne. Chez d'autres thérapsides comme Thrinaxodon, Bauria et Olivierosuchus, la branche nasale externe se divise généralement en trois branches ou plus. Tous ces canaux auraient amené des nerfs et des tissus riches en nutriments aux racines des canines et au reste de la mâchoire supérieure[4],[13].
Classification
En , Euchambersia est attribué à la famille des Euchambersiidae, alors nouvellement nommée par Lieuwe Dirk Boonstra[14],[15]. Celui-ci, lors de sa description, orthographie mal le taxon en « Euchambersidae », qui est un latin impropre. Par la suite, ce nom est corrigé par Friedrich von Huene en . Les Euchambersiidae ont été initialement considérées comme séparées des familles des Moschorhinidae et des Annatherapsididae. En , Christiane Mendrez reclasse ces groupes en tant que sous-familles étroitement apparentées, renommés Annatherapsidinae, Moschorhininae et Euchambersiinae au sein du groupe plus large et redéfini des Moschorhinidae (bien qu'elle continue d'utiliser le nom Annatherapsididae)[16].
L'analyse phylogénétique de James Hopson et Herb Barghusen en soutient l'hypothèse de Christiane Mendrez de trois sous-familles au sein des Moschorhinidae, mais les deux paléontologues préfèrent utiliser le nom Euchambersiidae comme synonyme. En , Adam Huttenlocker et ses collègues font valoir que les noms Annatherapsididae, Moschorhinidae et Euchambersiidae sont des synonymes juniors d'Akidnognathidae, puisque le genre Akidnognathus est le premier à avoir été décrit au sein de ce taxon[16]. Ce nom atteint par la suite une acceptation plus large parmi les chercheurs[16],[17],[18]. Huttenlocker et Christian Sidor (en) redéfinissent plus tard les Moschorhininae comme faisant partie des Akidnognathidae, à l'exception des genres Annatherapsidus et Akidnognathus[19].


En , Mikhaïl Ivakhnenko inclut les Akidnognathidae (alors désigné sous le synonyme Euchambersiidae dans son article) en tant que groupe frère de la famille des Whaitsiidae au sein de la super-famille des Whaitsioidea[15]. Cependant, d'autres chercheurs n'incluent pas les Akidnognathidae dans ce taxon. Les analyses phylogénétiques d'Huttenlocker et Sidor découvrent que les Akidnognathidae sont plutôt proches des Chthonosauridae, les deux formant le groupe frère d'un clade contenant les Whaitsioidea et les Baurioidea. La topologie récupérée par l'analyse de d'Huttenlocker et Sidor est montré ci-dessous[19] :
| ◄ Therocephalia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paléobiologie

Les grandes fosses situés dans les os maxillaires d'Euchambersia font l'objet d'un débat permanent quant à leurs fonctions. La plupart des chercheurs s'accordent pour dire qu'elles contenaient une sorte de glande sécrétoire. Broom soutenait initialement l'idée que les fosses auraient contenu des glandes salivaires parotides[1], une proposition qui sera plus tard rejetée par Boonstra et Jean-Pierre Lehman, qui notent que les glandes parotides ont tendance à être placées derrière l'œil[14],[20]. Boonstra suggère respectivement que les fosses aurait contenu des glandes lacrymales[14], tandis que Lehman propose que ces dernières contiendrait des glandes de Harder[20]. Cependant, l'hypothèse de Lehman s'avère être peu probable, car les glandes de Harder sont généralement placées à l'intérieur de l'orbite[4]. Franz Nopcsa émet la théorie que les fosses dans les maxillaires auraient abrité des glandes à venin, qui peuvent être dérivées de glandes lacrymales, les canines striées et les encoches derrière les canines ayant permis au venin de s'écouler passivement dans la circulation sanguine de la proie[21]. Cette hypothèse a été largement acceptée tout au long du XXe siècle[17],[22],[23],[24] et la morphologie caractéristique d'Euchambersia a été utilisée pour soutenir d'éventuelles adaptations venimeuses parmi divers autres animaux préhistoriques[9],[25],[26], incluant les thérocéphales apparentés Megawhaitsia et Ichibengops[15],[27].

Une grande partie de cette acceptation est basée sur l'hypothèse erronée que les canines sont rainurées au lieu d'être striées[3], les canines rainurées d'Euchambersia seraient parallèles aux crocs de divers serpents venimeux ainsi qu'aux incisives libérant du venin des solénodontes actuels[24]. Cette interprétation, apparue constamment dans la littérature scientifique publiée après , est déterminée par Julien Benoit comme étant le résultat de la propagation du diagramme trop reconstruit du crâne de Broom, sans le contexte des spécimens réels. Cette ligne de preuve est soulevée pour soutenir la nécessité d'une réévaluation de l'hypothèse d'une morsure venimeuse d'Euchambersia[3]. De plus, Benoit fait valoir que les canines cannelées et striées ne sont pas nécessairement associées aux animaux venimeux non plus, comme le montre leur présence chez les hippopotames, les muntjacs et les babouins, chez lesquels elles jouent un rôle pour le toilettage ou l'aiguisage des dents[24],[3],[28]. Chez les deux derniers, les canines striées sont également accompagnées d'une fosse devant l'œil, qui n'a aucun lien avec le venin[24],[29]. De plus, les dents rainurées et striées des serpents non venimeux sont utilisées pour réduire la traînée d'aspiration lors de la capture de proies glissantes comme les poissons ou les invertébrés[30].
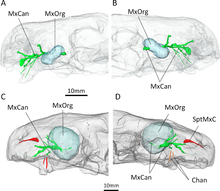
La tomodensitométrie des spécimens connus d'Euchambersia par Benoit et al. est plus tard utilisée pour fournir un support plus concret en faveur de l'hypothèse de la possible venimosité. Les canaux menant vers et depuis les fosses maxillaires, comme le révèlent les analyses, auraient principalement soutenu le nerf trijumeau ainsi que les vaisseaux sanguins[31],[32],[33]. Cependant, le fait que les canaux conduisent également et directement à la racine des canines suggère qu'ils auraient eu un rôle secondaire dans l'apport de venin. En tout, Euchambersia semble avoir eu une glande à venin, logée dans les fosses des maxillaires, un mécanisme de délivrance du venin, les canaux des maxillaires, et un instrument par lequel une blessure pour la livraison de venin peut être infligée, à savoir les canines striées, qui répondent aux critères d'un animal venimeux tel que défini par Wolfgang Bücherl[34]. Benoit et al. notent que cela ne démontre pas de manière concluante la venimosité d'Euchambersia, en particulier compte tenu des objections précédemment énoncées. De plus, aucun animal vivant n'est doté d'un système de distribution analogue au système proposé pour Euchambersia[N 1].
Une autre hypothèse suggérée par Benoit et al. implique une sorte d'organe sensoriel occupant les fosses dans les maxillaires. Uniquement parmi les thérapsides[13], le canal à l'intérieur du maxillaire est exposé à l'arrière de la fosse maxillaire, ce qui implique que le canal, portant le nerf trijumeau, a pu s'étendre à travers la fosse, en dehors du contour du crâne. Benoit et al. émettent donc l'hypothèse que la fosse peut avoir soutenu un organe sensoriel spécialisé, analogue à l'organe des crotales et d'autres serpents actuels, ou alternativement un ganglion de cellules nerveuses[38]. Il est également possible que cet organe remplace l'œil pariétal chez Euchambersia, comme le fait l'organe de la fosse des vipères[39]. Cependant, un tel organe sensoriel élargi serait non précédés chez les tétrapodes du Permien, et les quelques autres thérocéphales auxquels manque également un œil pariétal n'ont pas non plus de fosses dans les maxillaires[40]. Ainsi, Benoit et al. considèrent l'hypothèse du venin comme étant la plus plausible[4]. En , l'officialisation de la découverte du spécimen bien conservé de la deuxième espèce, E. liuyudongi, prouvent que ni le museau ni l'orbite montre des signes d'un glande venimeuse. Seules les glandes préorbitales, c'est à dire odorantes, ont été trouvées, soutenant l'hypothèse de la « glande odorante », bien que des tomodensitogrammes soient nécessaires pour plus de connaissances sur la dentition et le crâne de la nouvelle espèce[7].
Paléoécologie
Euchambersia mirabilis

D'après les archives fossiles, la zone d'assemblage supérieure de Cistecephalus, lieu d'où E. mirabilis est connu, correspond à une plaine inondable couvertes de nombreux petits ruisseaux formant des méandres. Le niveau d'eau dans ces fleuves dépendait probablement des saisons[5]. À en juger par le pollen conservé dans cette zone, le taxon pollinique Pityosporites (qui provient probablement d'une plante similaire à Glossopteris) est très commun, formant environ 80 à 90 % du pollen découvert, bien que les sédiments prédominants ne sont pas idéaux pour la conservation du pollen[41].
Au sein même de cette zone, de nombreux thérocéphales contemporains d'Euchambersia ont été identifiés, tels que Hofmeyria (en), Homodontosaurus (en), Ictidostoma (en), Ictidosuchoides (en), Ictidosuchops (en), Macroscelesaurus (en), Polycynodon (en) et Proalopecopsis. Des gorgonopsiens tels que Aelurognathus, Aelurosaurus, Aloposaurus, Arctognathus, Arctops, Cerdorhinus (en), Clelandina, Cyonosaurus, Dinogorgon, Gorgonops, Lycaenops, Rubidgea, Scylacops et Sycosaurus sont aussi présents[5].
Un des herbivores les plus abondant dans la zone d'assemblage de Cistecephalus est le dicynodonte Diictodon, avec plus de 1 900 spécimens connus[N 2]. D'autres dicynodontes découverts sont Aulacephalodon (en), Cistecephalus, Dicynodon, Dicynodontoides (en), Digalodon (en), Dinanomodon (en), Emydops, Endothiodon (en), Kitchinganomodon (en), Oudenodon (en), Palemydops (en), Pelanomodon (en), Pristerodon (en) et Rhachiocephalus (en). Des biarmosuchiens comme Lemurosaurus (en), Lycaenodon (en), Paraburnetia (en) et Rubidgina (en) ou encore des cynodontes non mammaliens comme Cynosaurus (en) et Procynosuchus sont connus de cette zone. D'autres tétrapodes non synapsides tels que des sauropsides incluant Younginia (en), Anthodon, Milleretta, Nanoparia (en), Owenetta (en) et Pareiasaurus ou des amphibiens temnospondyles comme Rhinesuchus sont également connus[5].
Euchambersia liuyudongi
La formation de Naobaogou en Chine, d'où E. liuyudongi a été découvert, est l'une des faunes du Permien supérieur les plus diversifié situé en dehors d'Afrique et de Russie, endroits où les tétrapodes de cette période sont majoritairement répandus[7]. Ce lieu préserve de nombreux fossiles de dicynodontes tels que Diictodon, Daqingshanodon (en) ou Turfanodon (en), des parareptiles comme Gansurhinus (en) et Elginia (en) et même des chroniosuchiens comme Laosuchus[7],[44],[45]. D'autres thérocéphales, incluant Caodeyao et des akidnognathidés proches du genre Euchambersia, tels Shiguaignathus ou Jiufengia, sont aussi connus au sein du site[7],[44].
La présence du genre Euchambersia dans cette formation est la première preuve d'une large distribution de thérocéphales en Chine et en Afrique du Sud. Cela renforce l'idée d'une distribution mondiale soutenue des thérapsides et illustre que des morphotypes disparates et assez inhabituels étaient représentés des deux côtés de la Pangée durant cette période[7].
Culture populaire
En raison de la découverte assez spéciale des crânes attribués à cet animal, Euchambersia est souvent représenté par plusieurs médias comme possédant du venin[3] :
- une meute de thérocéphales semblable à Euchambersia apparaît dans la septième séquence du documentaire Sur la terre des géants, dans laquelle l'un d'entre eux chasse un Lystrosaurus en utilisant une attaque venimeuse[3],[46] ;
- des grands thérocéphales apparaissent dans le quatrième épisode de la quatrième saison de la série Nick Cutter et les Portes du temps, où il est également présenté comme étant venimeux, bien que mal reconstitué et présenté avec une taille très exagérée[46].
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Euchambersia » (voir la liste des auteurs).
Références
- (en) R. Broom, « Notices of some new genera and species of Karroo fossil reptiles », Records of the Albany Museum, vol. 4, no 1, , p. 161-166.
- (en) James W. Kitching, The Distribution of the Karroo Vertebrate Fauna: With Special Reference to Certain Genera and the Bearing of this Distribution on the Zoning of the Beaufort Beds, Johannesburg, Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, , 1-131 p. (ISBN 9780854944279)
- (fr + en) J. Benoit, « Revue de l’hypothèse du “thérocéphalien venimeux” et de la façon dont les représentations successives de Euchambersia ont influencé son succès et vice versa », Bulletin de la Société géologique de France, vol. 187, no 4, , p. 217-224 (DOI 10.2113/gssgfbull.187.4-5.217, S2CID 88595137, lire en ligne).
- (en) J. Benoit, L.A. Norton, P.R. Manger et B.S. Rubidge, « Reappraisal of the envenoming capacity of Euchambersia mirabilis (Therapsida, Therocephalia) using μCT-scanning techniques », PLoS ONE, vol. 12, no 2, , e0172047 (PMID 28187210, PMCID 5302418, DOI 10.1371/journal.pone.0172047
 , Bibcode 2017PLoSO..1272047B).
, Bibcode 2017PLoSO..1272047B). - (en) R. Smith, B. Rubidge et Merrill van der Walt, Forerunners of Mammals: Radiation, Histology, Biology, Bloomington, Indiana University Press, , 31-64 p. (ISBN 978-0-253-00533-5, lire en ligne), « Therapsid Biodiversity Patterns and Palaeoenvironments of the Karoo Basin, South Africa ».
- (en) B.S. Rubidge, D.H. Erwin, J. Ramezani, S.A. Bowring et W.J. de Klerk, « High-precision temporal calibration of Late Permian vertebrate biostratigraphy: U-Pb zircon constraints from the Karoo Supergroup, South Africa », Geology, vol. 41, no 3, , p. 363-366 (DOI 10.1130/G33622.1, Bibcode 2013Geo....41..363R, S2CID 128626661, lire en ligne).
- (en) J. Liu et F. Abdala, « The emblematic South African therocephalian Euchambersia in China: a new link in the dispersal of late Permian vertebrates across Pangea », Biology Letters, vol. 18, no 7, , p. 20220222 (ISSN 1744-957X, PMID 35857894, PMCID 9278400, DOI 10.1098/rsbl.2022.0222 )
- (en) K. Andersson, D. Norman et L. Werdelin, « Sabretoothed Carnivores and the Killing of Large Prey », PLOS ONE, vol. 6, no 10, , e24971 (PMID 22039403, PMCID 3198467, DOI 10.1371/journal.pone.0024971 , Bibcode 2011PLoSO...624971A).
- (en) H.-D. Sues, « Venom-conducting teeth in a Triassic reptile », Nature, vol. 351, no 6322, , p. 141-143 (DOI 10.1038/351141a0, Bibcode 1991Natur.351..141S, S2CID 4306912, lire en ligne)
- (en) D.W. Kermack et K.A. Kermack, The Evolution of Mammalian Characters, Springer US, , 66-68 p. (ISBN 978-1-4684-7819-8, DOI 10.1007/978-1-4684-7817-4, lire en ligne), « Dentitions, Tooth-Replacement and Jaw Articulation ».
- (en) K.A. Kermack, « Tooth Replacement in Mammal-Like Reptiles of the Suborders Gorgonopsia and Therocephalia », Philosophical Transactions of the Royal Society B, vol. 240, no 670, , p. 95-133 (DOI 10.1098/rstb.1956.0013, Bibcode 1956RSPTB.240...95K).
- (en) J. A. Hopson, « Tooth replacement in cynodont, dicynodont, and therocephalian reptiles », Journal of Zoology, vol. 142, no 4, , p. 625-654 (DOI 10.1111/j.1469-7998.1964.tb04632.x).
- (en) J. Benoit, P. R. Manger et B. S. Rubidge, « Palaeoneurological clues to the evolution of defining mammalian soft tissue traits », Scientific Reports, vol. 6, , p. 25604 (PMID 27157809, PMCID 4860582, DOI 10.1038/srep25604 , Bibcode 2016NatSR...625604B).
- (en) Boonstra L.D., « A contribution to the morphology of the mammal-like reptiles of the suborder Therocephalia », Annals of the South African Museum, vol. 31, , p. 215-267.
- (en) M. F. Ivakhnenko, « The First Whaitsiid (Therocephalia, Theromorpha) », Paleontological Journal, vol. 42, no 4, , p. 409-413 (DOI 10.1134/S0031030108040102, S2CID 140547244).
- (en) A. Huttenlocker, « An investigation into the cladistic relationships and monophyly of therocephalian therapsids (Amniota: Synapsida) », Zoological Journal of the Linnean Society, vol. 157, no 4, , p. 865-891 (DOI 10.1111/j.1096-3642.2009.00538.x , S2CID 84603632).
- (en) B. S. Rubidge et C. A. Sidor, « Evolutionary Patterns Among Permo-Triassic Therapsids », Annual Review of Ecology and Systematics, vol. 32, , p. 449-480 (DOI 10.1146/annurev.ecolsys.32.081501.114113, S2CID 51563288, lire en ligne [PDF]).
- (en) T. Sigurdsen, « New features of the snout and orbit of a therocephalian therapsid from South Africa », Acta Palaeontologica Polonica, vol. 51, no 1, , p. 63-75 (résumé, lire en ligne [PDF]).
- (en) A.K. Huttenlocker et C.A. Sidor, « The first karenitid (Therapsida, Therocephalia) from the upper Permian of Gondwana and the biogeography of Permo-Triassic therocephalians », Journal of Vertebrate Paleontology, vol. 36, no 4, , e1111897 (DOI 10.1080/02724634.2016.1111897, JSTOR 24740259, S2CID 130994874).
- (fr + en) J. Piveteau, Traité de Paléontologie. L'origine des mammifères et les aspects fondamentaux de leur évolution., t. 6, Paris, Elsevier Masson, (OCLC 637017332, DOI 10.1126/science.129.3355.1020.b, S2CID 239782868), « Therocephalia », p. 224-245
- (en) F. Nopcsa, « On the biology of the theromorphous reptile Euchambersia », Annals and Magazine of Natural History, 10e série, vol. 12, no 67, , p. 125-126 (DOI 10.1080/00222933308673757, S2CID 86174437).
- (en) D. M. S. Watson et A. S. Romer, « A classification of therapsid reptiles », Bulletin of the Museum of Comparative Zoology, vol. 114, , p. 35-89 (lire en ligne)
- (en) L. Van Valen, « Therapsids as Mammals », Evolution, vol. 14, no 3, , p. 304-313 (DOI 10.2307/2405973 , JSTOR 2405973, S2CID 88232897, lire en ligne [PDF])
- (en) K. E. Folinsbee, J. Muller et R. R. Reisz, « Canine Grooves: Morphology, Function, and Relevance to Venom », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 547-551 (DOI 10.1671/0272-4634(2007)27[547:cgmfar]2.0.co;2, JSTOR 30126324, S2CID 54602365, lire en ligne [PDF])
- (en) H.-D. Sues, « A reptilian tooth with apparent venom canals from the Chinle Group (Upper Triassic) of Arizona », Journal of Vertebrate Paleontology, vol. 16, no 3, , p. 571-572 (DOI 10.1080/02724634.1996.10011340, S2CID 129631972)
- (en) E. Gong, L. D. Martin, D. A. Burnham et A. R. Falk, « The birdlike raptor Sinornithosaurus was venomous », Proceedings of the National Academy of Sciences of the United States of America, vol. 107, no 2, , p. 766-768 (PMID 20080749, PMCID 2818910, DOI 10.1073/pnas.0912360107 , Bibcode 2010PNAS..107..766G)
- (en) Adam K. Huttenlocker, Christian A. Sidor et Kenneth D. Angielczyk, « A new eutherocephalian (Therapsida, Therocephalia) from the upper Permian Madumabisa Mudstone Formation (Luangwa Basin) of Zambia », Journal of Vertebrate Paleontology, vol. 35, no 5, , e969400 (DOI 10.1080/02724634.2015.969400, S2CID 83554630)
- (en) J. S. Mitchell, A. B. Heckert et H.-D. Sues, « Grooves to tubes: evolution of the venom delivery system in a Late Triassic "reptile" », Naturwissenschaften, vol. 97, no 12, , p. 1117-1121 (PMID 21060984, DOI 10.1007/s00114-010-0729-0, Bibcode 2010NW.....97.1117M, S2CID 10093308, lire en ligne [PDF]).
- (en) C. M. Orr, L. K. Delezene, J. E. Scott, M. W. Tocheri et G. T. Schwartz, « The comparative method and the inference of venom-delivery systems in fossil mammals », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 541-546 (DOI 10.1671/0272-4634(2007)27[541:TCMATI]2.0.CO;2, S2CID 45645935, lire en ligne [PDF]).
- (en) R. H. Vaeth, D. A. Rossman et W. Shoop, « Observations of Tooth Surface Morphology in Snakes », Journal of Herpetology, vol. 19, no 1, , p. 20-26 (DOI 10.2307/1564416, JSTOR 1564416, S2CID 87793935).
- (en) A. D'A. Bellairs, « Observations on the snout of Varanus, and a comparison with that of other lizards and snakes », Journal of Anatomy, vol. 83, no 2, , p. 116-146 (PMID 17105074, PMCID 1273152, lire en ligne [PDF]).
- (en) T. G. Abdel-Kader, R. S. Ali et N. M. Ibrahim, « The Cranial Nerves of Mabuya quinquetaeniata III: Nervus Trigeminus », Life Science Journal, vol. 8, no 4, , p. 650-669 (lire en ligne [PDF]).
- (en) D. B. Leitch et K. C. Catania, « Structure, innervation and response properties of integumentary sensory organs in crocodilians », Journal of Experimental Biology, vol. 215, no 23, , p. 4217-4230 (PMID 23136155, PMCID 4074209, DOI 10.1242/jeb.076836 ).
- (en) W. Bücherl, « Introduction », dans W. Bücherl, E. E. Buckley et V. Deulofeu, Venomous Animals and their Venoms, vol. 1, New York, Academic Press, , 732 p. (ISBN 978-1-483-22949-2, DOI 10.1016/B978-1-4832-2949-2.50006-0, S2CID 86826264, résumé), p. 9-12
- (en) B. G. Fry et al., « A central role for venom in predation by Varanus komodoensis (Komodo Dragon) and the extinct giant Varanus (Megalania) priscus », Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no 22, , p. 8969-8974 (PMID 19451641, PMCID 2690028, DOI 10.1073/pnas.0810883106 , Bibcode 2009PNAS..106.8969F).
- (en) R. Ligabue-Braun, H. Verli et C. R. Carlini, « Venomous mammals: A review », Toxicon, vol. 59, no 7, , p. 680-695 (PMID 22410495, DOI 10.1016/j.toxicon.2012.02.012, S2CID 5055413).
- (en) S. A. Weinstein, T. L. Smith et K. V. Kardong, « Reptile Venom Glands: Form, Function, and Future », dans S. P. Mackessy,, Handbook of Venoms and Toxins of Reptiles, Boca Raton, CRC Press, , PDF (lire en ligne), p. 65–91
- (en) R. C. Goris, « Infrared Organs of Snakes: An Integral Part of Vision », Journal of Herpetology, vol. 45, no 1, , p. 2-14 (DOI 10.1670/10-238.1, S2CID 86066152).
- (en) A. R. Krochmal, G. S. Bakken et T. J. LaDuc, « Heat in evolution's kitchen: evolutionary perspectives on the functions and origin of the facial pit of pitvipers (Viperidae: Crotalinae) », Journal of Experimental Biology, vol. 207, no 24, , p. 4231-4238 (PMID 15531644, DOI 10.1242/jeb.01278 , S2CID 13925793).
- (en) J. Benoit, F. Abdala, P. R. Manger et B. S. Rubidge, « The Sixth Sense in Mammalian Forerunners: Variability of the Parietal Foramen and the Evolution of the Pineal Eye in South African Permo-Triassic Eutheriodont Therapsids », Acta Palaeontologica Polonica, vol. 61, no 4, , p. 777-789 (DOI 10.4202/app.00219.2015 , S2CID 59143925).
- (en) J.M. Anderson, A Review of Gondwana Permian Palynology with Particular Reference to the Northern Karoo Basin of South Africa, vol. 41, coll. « Memoirs of the Botanical Survey of South Africa », , 42-58 p. (lire en ligne), « The microfloral succession: conclusions and discussion ».
- (en) R.M.H. Smith, B.S. Rubidge, M.O. Day et J. Botha, « Introduction to the tetrapod biozonation of the Karoo Supergroup », South African Journal of Geology, vol. 123, no 2, , p. 131–140 (ISSN 1996-8590 et 1012-0750, DOI 10.25131/sajg.123.0009).
- (en) M.O. Day et R.M.H. Smith, « Biostratigraphy of the Endothiodon Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa », South African Journal of Geology, vol. 123, no 2, , p. 165–180 (ISSN 1996-8590 et 1012-0750, DOI 10.25131/sajg.123.0011, S2CID 225834576).
- (en) Jun Liu, « The tetrapod fauna of the upper Permian Naobaogou Formation of China: 6. Turfanodon jiufengensis sp. nov. (Dicynodontia) », PeerJ, vol. 9, , e10854 (PMID 33643709, PMCID 7896508, DOI 10.7717/peerj.10854 )
- (en) J. Liu et J. Chen, « The tetrapod fauna of the upper Permian Naobaogou Formation of China: 7. Laosuchus hun sp. nov. (Chroniosuchia) and interrelationships of chroniosuchians », Journal of Systematic Palaeontology, vol. 18, no 24, , p. 2043-2058 (DOI 10.1080/14772019.2021.1873435, S2CID 232116225)
- (en) « Study of the Euchambersia localities », sur sahris.sahra.org.za (consulté le ).
Voir aussi
Articles connexes
Vidéos
- (en) [vidéo] Our Bizarre, Possibly Venomous, Relative sur YouTube.
Liens externes
- Ressources relatives au vivant :
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’Afrique du Sud
Portail de l’Afrique du Sud  Portail de la Chine
Portail de la Chine
На других языках
[es] Euchambersia
Euchambersia es un género extinto de sinápsidos no mamíferos terápsidos terocéfalos que vivieron del Pérmico Superior - Triásico Inferior, hace unos 250 millones de años en lo que ahora es Sudáfrica. Sus restos fósiles, dos especímenes con cráneo y esqueleto postcraneal incompleto, aparecieron en el Karoo, provincia del Cabo Occidental.- [fr] Euchambersia
[ru] Эучамберсии
Эучамберсии[1] (лат. Euchambersia) — род хищных тероцефалов позднепермской эпохи, близкий к мосхорину, териогнату и аннатерапсиду, включающий два вида: Euchambersia mirabilis и Euchambersia liuyudongi[2].Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии