bio.wikisort.org - Animal
Cynodontes
Ne doit pas être confondu avec Cynodontidae.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Clade | Eutheriodontia |
Sous-ordre ou Clade
Owen, 1861
Taxons de rang inférieur
- †Charassognathidae
- †Procynosuchidae
- †Dviniidae
- †Madysauridae
- Epicynodontia (inclut les mammifères)
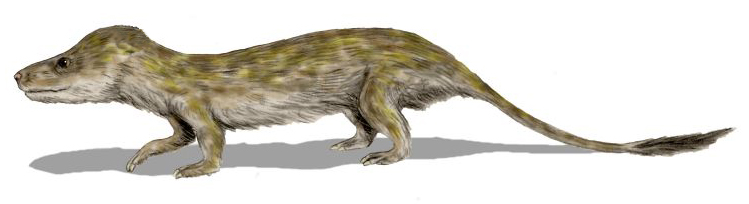
Les cynodontes (Cynodontia) forment un sous-ordre et un clade de thérapsides euthériodontes qui apparaissent durant le Permien supérieur, vers 260 millions d'années avant notre ère, et qui comprennent les mammifères (dont l'homme), leurs ancêtres et les groupes apparentés aujourd'hui éteints.
Ce groupe partage un ancêtre commun avec les thérocéphales et, avec ces derniers et les dicynodontes (Anomodontia), figure parmi les rares groupes de synapsides survivant à l'extinction Permien-Trias et qui recolonisent les niches écologiques délaissées par les gorgonopsiens lors de leur disparition, en concurrence avec les premiers dinosaures qui commencent à apparaître peu de temps après. Au fil des millions d'années, ils évoluent en plusieurs lignées d'herbivores et de carnivores.
Caractéristiques

Les premiers cynodontes[note 1] ont de nombreuses caractéristiques squelettiques des mammifères. Les dents sont complètement différenciées et la boîte crânienne renflée à l'arrière de la tête. En dehors de certains espèce du groupe-couronne des mammifères (notamment les theriéns), tous les cynodontes aurait probablement pondu des œufs. Les fenêtres temporales sont beaucoup plus grandes que celles de leurs ancêtres, et l'élargissement de l'arc zygomatique dans un crâne plus proche des mammifères permet une musculature de la mâchoire plus robuste. Ils ont aussi un palais secondaire qui manque chez les autres thérapsides plus primitifs, à l'exception des thérocéphales, qui sont les plus proches parents de ces derniers. Cependant, le palais secondaire des cynodontes comprend principalement les maxillaires et les palatins comme chez les mammifères, tandis que le palais secondaire des thérocéphales comprend principalement les maxillaires et le vomer. Le dentaire est le plus gros os de leurs mâchoires inférieures.
Les cynodontes possèdent probablement une certaine forme de métabolisme à sang chaud, c'est d'ailleurs pour cette raison qu'il y a de nombreuses reconstitutions de cynodontes comme ayant de la fourrure. Étant endothermiques, ils en ont peut-être besoin pour la thermorégulation, mais les preuves fossiles de leur fourrure (ou de leur absence) sont insaisissables. Les mammifères modernes ont des glandes de Harder sécrétant des lipides pour recouvrir leurs fourrures, mais l'empreinte révélatrice de cette structure ne se trouve qu'à partir du mammaliaforme Morganucodon et d'autres[1]. Néanmoins, des études récentes sur les coprolithes de thérapsides du Permien montrent que des thérapsides plus basaux peuvent avoir eu de la fourrure[2].
Les premiers cynodontes ont de nombreux petits foramens sur les os de leurs museaux, semblables à ceux des reptiles. Cela suggère qu'ils auraient eu des lèvres immobiles et non musclées comme celles des lézards, manquant de joues musclées[3]. Comme un visage musclé et mobile est nécessaire pour effectuer des mouvements de fouet et pour éviter d'endommager les vibrisses, il est peu probable que les premiers cynodontes possèdent ces derniers[4],[5]. Chez les cynodontes prozostrodontiens, le groupe qui comprend les mammifères, les foramens sont remplacés par un seul grand foramen infra-orbitaire, ce qui indique que le visage devient musclé et que des vibrisses auraient été présentes.
Les cynodontes dérivés disposent des os épipubiens, ceux-ci servent à renforcer le torse et à soutenir la musculature abdominale et des membres postérieurs, les aidant à développer une démarche dressée, mais au détriment d'une grossesse prolongée, forçant ces animaux à donner naissance à des jeunes hautement nidicoles comme chez les marsupiaux et les monotrèmes modernes. Seuls les placentaires, et peut-être Megazostrodon ou Erythrotherium, les perdent[6],[7]. Un spécimen de Kayentatherium démontre en effet qu'au moins les tritylodontidés (fin du trias) auraient déjà un style de reproduction fondamentalement semblable à celui des marsupiaux, mais produiraient des portées beaucoup plus élevées à environ 38 périnates ou peut-être des œufs[8].
Les cynodontes sont la seule lignée de synapsides connue à avoir des locomoteurs aériens, ce qui est identifié chez les haramiyidiens[9] et certains groupes de mammifères.
Le plus grand cynodonte non-mammalien connu à ce jour est Scalenodontoides, un traversodontidé, dont la longueur maximale du crâne est estimée à environ 617 millimètres sur la base d'un spécimen fragmentaire[10].
Histoire évolutive


Les plus proches parents des cynodontes sont les thérocéphales, avec lesquels ils forment le clade Eutheriodontia[11].
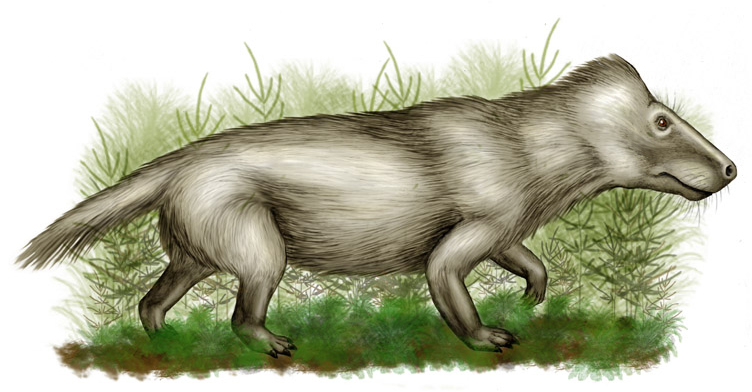
Les premiers cynodontes viennent des sédiments datant du Lopingien inférieur (Wuchiapingien inférieur) connus des zones d'assemblages en Afrique du Sud, appartenant à la famille basale des Charassognathidae. Les fossiles de cynodontes du Permien sont relativement rares en dehors de ce territoire, le genre le plus répandu étant Procynosuchus, connu en Afrique du Sud, en Allemagne, en Tanzanie, en Zambie et peut-être même de Russie[12].
Les cynodontes se développent rapidement en diversité après l'extinction Permien-Trias. Après le Trias inférieur, ils sont principalement représentés par les membres du clade avancé Eucynodontia, qui a deux subdivisions principales, les Cynognathia, à prédominance herbivore, et les Probainognathia, à prédominance carnivore. Au cours du Trias inférieur et moyen, la diversité des cynodontes est dominée par des membres des Cynognathia, les membres de Probainognathia ne devenant importants qu'à partir du Trias supérieur (début du Norien)[13]. Presque tous les cynodontes du Trias moyen sont connus du Gondwana, avec un seul genre trouvé dans l'hémisphère nord (Nanogomphodon). Parmi les groupes les plus dominants de cynodontes du Trias moyen et supérieur se trouvent les Traversodontidae, herbivores, principalement au Gondwana, qui atteignent un pic de diversité au Trias supérieur. Les mammifères sont originaires des cynodontes probainognathiens datant de la même période. Seuls deux groupes de cynodontes non-mammaliens existent au-delà de la fin du Trias, tous deux appartenant au même groupe. Le premier forme les insectivores de la famille des Tritheledontidae, qui a duré brièvement jusqu'au Jurassique inférieur. Le second forme les herbivores de la famille des Tritylodontidae, qui apparaissent durant le Trias supérieur, qui sont abondants et diversifiés au cours du Jurassique, principalement dans l'hémisphère nord, jusqu'au Crétacé inférieur en Asie[14]. Le tritylodontidé le plus récent connu est Fossiomanus de la formation de Jiufotang, en Chine, vieille de 120 millions d'années[15].
Au cours de leur évolution, le nombre d'os de la mâchoire des cynodontes diminue. Cette évolution vers un os unique pour la mandibule ouvre la voie à d'autres os de la mâchoire, l'os articulaire et l'angulaire, pour migrer vers le crâne, où ils font partie du système auditif des mammifères.
Les cynodontes possèdent également un palais secondaire situé au-dessus de la cavité buccale. Cela amene le flux d'air des narines à se déplacer vers l'arrière de la bouche au lieu de passer directement à travers elle, permettant aux cynodontes de mâcher et de respirer en même temps. Cette caractéristique est toujours présente chez les mammifères.
Répartition
Les fossiles de cynodontes sont trouvables en Amérique du Sud, en Afrique, en Inde, en Antarctique[16], en Asie[17], en Europe[18] et en Amérique du Nord[19].
Taxinomie
Historique
Richard Owen décrit le taxon Cynodontia en 1861, qu'il attribue à Anomodontia en tant que famille[20]. Robert Broom (1913) reclasse Cynodontia comme un infra-ordre, depuis retenu par d'autres, dont Colbert & Kitching (1977), Carroll (1988), Gauthier et al. (1989) ainsi que Rubidge & Christian Sidor (2001)[21]. Olson (1966), Colbert & Kitching (1977) classent Cynodontia au sein du taxon Theriodontia, et Rubridge et Sidor (2001) dans le sous-groupe Eutheriodontia. William King Gregory (1910), Broom (1913), Carroll (1988), Gauthier et al. (1989), Hopson & Kitching (2001) et Botha et al.(2007) considèrent Cynodontia comme appartenant à Therapsida. Botha et al. (2007) semble suivre Owen (1861), mais sans préciser le rang taxonomique[22],[23].
Phylogénie



Ci-dessous, un cladogramme de Ruta, Botha-Brink, Mitchell et Benton (2013) montrant une hypothèse de relations entre les cynodontes[13] :
| Cynodontia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| → †Cynognathia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| → Probainognathia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cynodontia dans Therapsida
Le cladogramme ci-dessous présente la phylogénie des thérapsides selon T. S. Kemp (2011)[24] basée selon la proposition d'Hopson et Barghausen (1986)[25] :
| ◄ Therapsida |
| |||||||||||||||||||||||||||||||||
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) J.A. Hopson et J.W. Kitching, « A probainognathian cynodont from South Africa and the phylogeny of non-mammalian cynodonts », Bull. Mus. Comp. Zool., vol. 156, , p. 5–35
- (en) Davis, Dwight (1961). "Origin of the Mammalian Feeding Mechanism". Am. Zoologist, 1:229–234.
Voir aussi
- Therapsida
- Eutheriodontia
- Therocephalia
- Histoire évolutive des mammifères
Liens externes
- Ressource relative au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Cynodont » (voir la liste des auteurs).
- En raison de leurs dentitions, le mot « cynodonte » veux littéralement dire « dent de chien » (du grec ancien κύων, κυνός / kúôn, kunós « chien » et ὀδούς, ὀδόντος / odoús, odóntos « dent »).
Références
- (en) J.A. Ruben et T.D. Jones, « Selective Factors Associated with the Origin of Fur and Feathers », American Zoologist, vol. 40, no 4, , p. 585–596 (DOI 10.1093/icb/40.4.585)
- (en) Piotr Bajdek, « Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia », Lethaia, vol. 49, no 4, , p. 455–477 (DOI 10.1111/let.12156)
- (en) Richard Estes, « Cranial anatomy of the cynodont reptile Thrinaxodon liorhinus », Bulletin Museum of Comparative Zoology, vol. 125, , p. 165–180
- (en) J. Benoit, P. R. Manger et B. S. Rubidge, « Palaeoneurological clues to the evolution of defining mammalian soft tissue traits », Scientific Reports, vol. 6, no 1, , p. 25604 (ISSN 2045-2322, PMCID 4860582, DOI 10.1038/srep25604, lire en ligne)
- (en) Julien Benoit, Irina Ruf, Juri A. Miyamae, Vincent Fernandez, Pablo Gusmão Rodrigues et Bruce S. Rubidge, « The Evolution of the Maxillary Canal in Probainognathia (Cynodontia, Synapsida): Reassessment of the Homology of the Infraorbital Foramen in Mammalian Ancestors », Journal of Mammalian Evolution, vol. 27, no 3, , p. 329–348 (ISSN 1064-7554, DOI 10.1007/s10914-019-09467-8, lire en ligne)
- (en) Michael L. Power, Jay Schulkin. The Evolution of the Human Placenta. p. 68–.
- (en) Jason A. Lillegraven, Zofia Kielan-Jaworowska, William A. Clemens, Mesozoic Mammals: The First Two-Thirds of Mammalian History, University of California Press, 17 December 1979 – 321
- (en) Eva A. Hoffman et Timothy B. Rowe, « Jurassic stem-mammal perinates and the origin of mammalian reproduction and growth », Nature, vol. 561, no 7721, , p. 104–108 (PMID 30158701, DOI 10.1038/s41586-018-0441-3, Bibcode 2018Natur.561..104H, S2CID 205570021)
- (en) Qing-Jin Meng, David M. Grossnickle, Liu Di, Yu-Guang Zhang, April I. Neander, Qiang Ji et Zhe-Xi Luo, « New gliding mammaliaforms from the Jurassic », Nature, vol. 548, no 7667, , p. 291–296 (PMID 28792929, DOI 10.1038/nature23476, Bibcode 2017Natur.548..291M, S2CID 205259206)
- (en) Frederick Tolchard, Christian F. Kammerer, Richard J. Butler, Christophe Hendrickx, Julien Benoit, Fernando Abdala et Jonah N. Choiniere, « A new large gomphodont from the Triassic of South Africa and its implications for Gondwanan biostratigraphy », Journal of Vertebrate Paleontology, , e1929265 (ISSN 0272-4634, DOI 10.1080/02724634.2021.1929265, lire en ligne)
- (en) Kenneth D. Angielczyk et Christian F. Kammerer, Mammalian Evolution, Diversity and Systematics, De Gruyter, , 117–198 p. (ISBN 978-3-11-034155-3, DOI 10.1515/9783110341553-005, S2CID 92370138, lire en ligne), « Non-Mammalian synapsids: the deep roots of the mammalian family tree »
- (en) Adam K. Huttenlocker et Christian A. Sidor, « A Basal Nonmammaliaform Cynodont from the Permian of Zambia and the Origins of Mammalian Endocranial and Postcranial Anatomy », Journal of Vertebrate Paleontology, vol. 40, no 5, , e1827413 (ISSN 0272-4634, DOI 10.1080/02724634.2020.1827413, lire en ligne)
- (en) M. Ruta, J. Botha-Brink, S. A. Mitchell et M. J. Benton, « The radiation of cynodonts and the ground plan of mammalian morphological diversity », Proceedings of the Royal Society B: Biological Sciences, vol. 280, no 1769, , p. 20131865 (PMID 23986112, PMCID 3768321, DOI 10.1098/rspb.2013.1865)
- (en) Fernando Abdala et Leandro C. Gaetano, The Late Triassic Record of Cynodonts: Time of Innovations in the Mammalian Lineage, vol. 46, Cham, Springer International Publishing, , 407–445 p. (ISBN 978-3-319-68008-8, DOI 10.1007/978-3-319-68009-5_11, lire en ligne)
- (en) Fangyuan Mao, Chi Zhang, Cunyu Liu et Jin Meng, « Fossoriality and evolutionary development in two Cretaceous mammaliamorphs », Nature, (ISSN 0028-0836, DOI 10.1038/s41586-021-03433-2, lire en ligne)
- (en) Fernando Abdala et Ana Maria Ribeiro, « Distribution and diversity patterns of Triassic cynodonts (Therapsida, Cynodontia) in Gondwana », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 286, nos 3–4, , p. 202–217 (DOI 10.1016/j.palaeo.2010.01.011, Bibcode 2010PPP...286..202A, lire en ligne)
- (en) Spencer G. Lucas, Chinese Fossil Vertebrates, New York, Chichester, West Sussex, Columbia University Press, (ISBN 0-231-08482-X, lire en ligne), p. 133
- (en) Nicholas C. Fraser et Hans-Dieter Sues, In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods, Cambridge University Press,
- (en) Hans-Dieter Sues, Paul E. Olsen et Joseph G. Carter, « A Late Triassic Traversodont Cynodont From the Newark Supergroup of North Carolina », Journal of Vertebrate Paleontology, vol. 19, no 2, , p. 351–354 (DOI 10.1080/02724634.1999.10011146, lire en ligne)
- (en) Classification of R. Owen 1861.
- (en) Classification of B. S. Rubidge and C. A. Sidor 2001
- (en) R. Broom. 1913. A revision of the reptiles of the Karroo. Annals of the South African Museum 7(6):361–366
- (en) S. H. Haughton and A. S. Brink. 1954. A bibliographical list of Reptilia from the Karroo Beds of South Africa. Palaeontologia Africana 2:1–187
- (en) T. S. Kemp, « The origin and radiation of therapsids », dans Chinsamy-Turan A., Forerunners of Mammals, Bloomington, Indiana University Press, (ISBN 978-0-253-35697-0, lire en ligne), p. 3-30.
- (en) J. A. Hopson et H. R. Barghusen, The Ecology and Biology of Mammal-Like Reptiles, Washington D. C., Smithsonian Institution Press, , 83-106 p. (OCLC 468801431), « An analysis of therapsid relationships »
 Portail de la paléontologie
Portail de la paléontologie  Portail origine et évolution du vivant
Portail origine et évolution du vivant  Portail des mammifères
Portail des mammifères
На других языках
[es] Cynodontia
Los cinodontos (Cynodontia, griego «dientes de perro») son un clado de terápsidos que aparecieron por primera vez en el Carbonífero, hace unos 300 millones de años. El grupo incluye a todos los modernos mamíferos, así como a sus antepasados extintos y a parientes cercanos. Los cinodontes no mamíferos se extendieron por todo el sur de Gondwana y están representados por fósiles de América del Sur, África, India y la Antártida. En los continentes del norte, los fósiles se han encontrado en el este de América del Norte, así como en Bélgica y el noroeste de Francia. Cynodontia es uno de los grupos más diversos de terápsidos.- [fr] Cynodontia
[ru] Цинодонты
Цинодо́нты[1] (лат. Cynodontia, буквально — «собакозубые») — клада терапсид в составе группы териодонтов. Все современные цинодонты относятся к млекопитающим (Mammalia)[2][3]. Ныне вымершие .mw-parser-output .ts-comment-commentedText{border-bottom:1px dotted;cursor:help}@media(hover:none){.mw-parser-output .ts-comment-commentedText:not(.rt-commentedText){border-bottom:0;cursor:auto}}немаммальные цинодонты, в том числе и предки млекопитающих, обладали почти всеми признаками своих продвинутых родственников и потомков (больше всего напоминали современных однопроходных)[4]. Зубы были полностью дифференцированы (их дифференциация продолжалась в течение всего периода существования). Немаммальные цинодонты откладывали яйца, подобно современным однопроходным млекопитающим и рептилиям. Скорее всего, они были теплокровными и покрытыми шерстью. Изучение строения черепа цинодонтов рода Dvinia показало, что у двинии присутствует разделение носовой полости на этмоидную и максиллярную турбиналии, что наблюдается только у теплокровных форм (птиц и млекопитающих) и связано с нагреванием вдыхаемого воздуха. Таким образом, имеются основания полагать, что теплокровность появилась уже на уровне пермских цинодонтов[5]. У вида Microdocodon gracilis, жившего около 165 млн лет назад, имелась седловидная подъязычная кость, очень похожая на подъязычную кость современных млекопитающих[6].Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии