bio.wikisort.org - Animalia
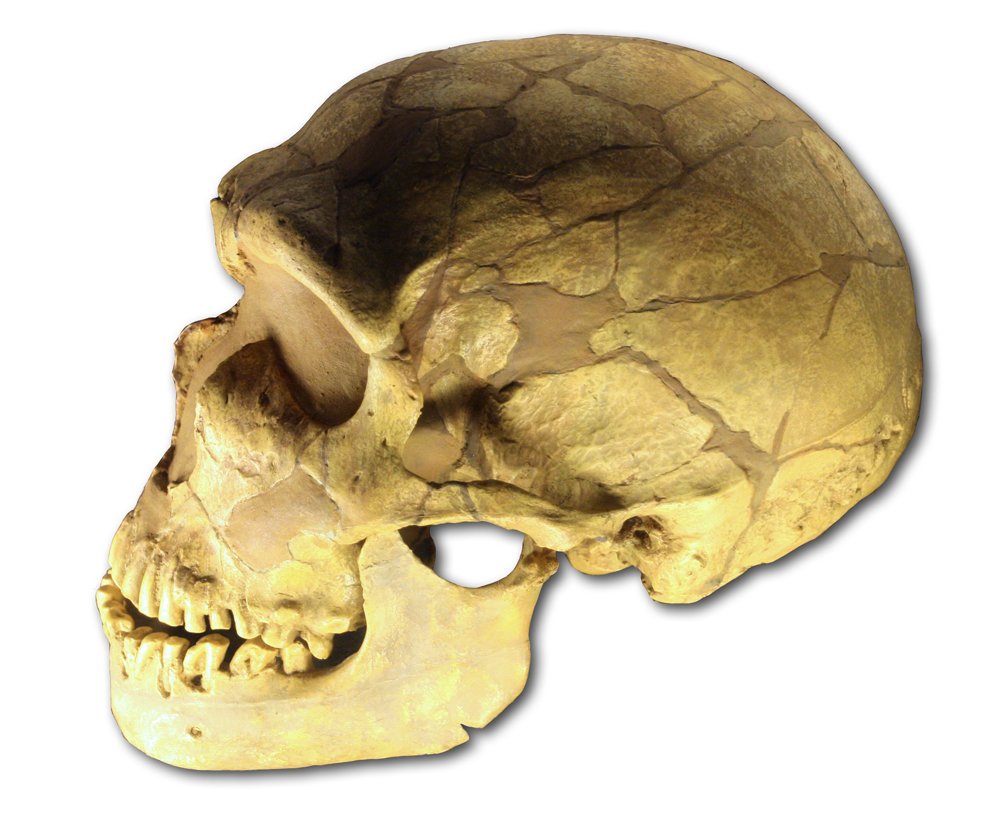
Homo neanderthalensis, King, 1864, comunemente detto uomo di Neandertal[1], è un ominide strettamente affine all'Homo sapiens che visse nel periodo paleolitico medio, compreso tra i 200 000 e i 40 000 anni fa.
| Uomo di Neandertal | |
|---|---|
 | |
| Stato di conservazione | |
Fossile Periodo di fossilizzazione: Pleistocene superiore (Paleolitico medio) | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Sottoregno | Eumetazoa |
| Superphylum | Deuterostomia |
| Phylum | Chordata |
| (clado) | Craniata |
| Subphylum | Vertebrata |
| Infraphylum | Gnathostomata |
| Superclasse | Tetrapoda |
| Classe | Mammalia |
| Sottoclasse | Theria |
| Infraclasse | Eutheria |
| Superordine | Euarchontoglires |
| (clado) | Euarchonta |
| Ordine | Primates |
| Sottordine | Haplorrhini |
| Infraordine | Simiiformes |
| Parvordine | Catarrhini |
| Superfamiglia | Hominoidea |
| Famiglia | Hominidae |
| Sottofamiglia | Homininae |
| Tribù | Hominini |
| Sottotribù | Hominina |
| Genere | Homo |
| Specie | H. neanderthalensis |
| Nomenclatura binomiale | |
| Homo neanderthalensis King, 1864 | |
| Sinonimi | |
|
Homo sapiens neanderthalensis | |
| Nomi comuni | |
|
Neanderthal, Neandertal, Paleantropo | |

Prende il nome dalla valle di Neander (Neandertal) presso Düsseldorf in Germania, dove vennero ritrovati i primi resti fossili. Fu un "Homo" molto evoluto, in possesso di tecnologie litiche elevate e dal comportamento sociale piuttosto avanzato, al pari dei sapiens di diversi periodi paleolitici.
Convissuto nell'ultimo periodo della sua esistenza con lo stesso Homo sapiens, l'Homo neanderthalensis scomparve in un tempo relativamente breve, evento che costituisce un enigma scientifico oggi attivamente studiato.
Storia
I resti che diedero il nome alla specie furono scoperti nell'agosto 1856 da scavatori di calcare in una grotta denominata "Kleine Feldhofer" nella valle di Neander, nei pressi della località di Feldhof in Germania. I reperti furono mostrati a Johann Fuhlrott, insegnante di scienze, che riconobbe trattarsi di una nuova specie del genere Homo. Della scoperta dei fossili venne dato annuncio ufficiale solo il 4 febbraio 1857[2].
Documentata fra 130 000 (per le forme arcaiche) e 30 000 (documentata con reperti fossili) −22 000 (in assenza di fossili ma con discusse prove culturali) anni fa principalmente in Europa e Asia, e limitatamente in Africa.
Alcuni studi del 2010 suggeriscono, tra alcune ipotesi probabili relative alla vicinanza genetica tra H. neanderthalensis e H. sapiens, che ibridazioni fra i due possano avere avuto luogo nel Vicino Oriente all'incirca tra 80 000 e 50 000 anni fa, per la presenza nell'uomo contemporaneo di una percentuale compresa tra l'1 e il 4% di materiale genetico specificamente neandertaliano. Tali tracce genetiche sono presenti negli eurasiatici e nei nativi americani, ma non negli africani subsahariani: ciò suggerisce, tra almeno quattro diverse ipotesi, che l'ibridazione possa avere avuto luogo nei primi stadi della migrazione della specie umana fuori dall'Africa, presumibilmente quando venne a contatto con i Neanderthal che vivevano nel Vicino Oriente, circa 80 000 anni or sono[3][4].
In passato, la specie era stata chiamata anche "uomo di Neanderthal", dall'originale nome specifico scientifico, e Homo sapiens neanderthalensis quando era ancora considerato sottospecie dell'Homo sapiens; queste denominazioni talora si riscontrano ancora oggi e così pure avviene in altre lingue. Il problema del nome come sottospecifico non è, al contrario del primo, meramente formale, ma riflette, come verrà esposto più avanti, diversi possibili cammini evolutivi e differenti gradi di reincrocio con i sapiens.
Sulla estinzione dell'H. neanderthalensis sono in elaborazione numerose teorie.

Sintesi dell'aspetto esteriore
L'aspetto fisico esteriore del neandertaliano classico, alla luce delle conoscenze attuali, notevolmente incrementatesi rispetto alle prime ipotesi ed estrapolazioni otto-novecentesche, è quello di un uomo di altezza medio-bassa (1,60 m) perfettamente eretto e muscolarmente molto robusto, con un rapporto superficie/volume inferiore a quello delle altre specie di uomini, il che comporta una minore dispersione di calore e quindi una maggiore resistenza alle basse temperature. La statura media, in base a quarantacinque ossa lunghe[5] di almeno quattordici maschi e sette femmine, era di circa 164–168 cm per i maschi e circa 152–156 cm per le femmine[6]. I campioni di ventisei esemplari studiati nel 2010 mostrano un peso medio di 77,6 kg per i maschi e 66,4 kg per le femmine[7].
La testa, rispetto ad un sapiens, risulta allungata antero-posteriormente, anche se si sovrappone in genere alla variabilità sapiens, ha un volume cerebrale di 1500 cm³ in media, del 10% superiore agli uomini attuali[8], e arcate sopraccigliari sporgenti. Ha uno spiccato prognatismo mascellare e il mento può essere sfuggente, per lo meno nei tipi arcaici. Con il passare del tempo, in alcune zone e verso la fine del Paleolitico, si diffonde un tipo più gracile e con un mento osseo più pronunciato, mentre gli zigomi sono molto meno accentuati e le arcate sopraccigliari al contrario più sporgenti.
Una tesi esposta nel 2006 e confermata nel 2007[9], basata su ricerche avanzate con tecniche di biologia molecolare, ipotizza che la specie, in Europa, abbia sviluppato individui di carnagione bianca con capelli rossi: il tipo di pigmentazione è in accordo con la scarsa irradiazione solare (ultravioletta) del territorio colonizzato, analogamente alla distribuzione geografica attuale della pigmentazione nei tipi umani[10]. Ciononostante, si è evidenziato come la variabilità genetica della popolazione neandertaliana[11][12] suggerisca una variabilità del fenotipo piuttosto ampia, analogamente a quella attuale di H. sapiens.
Recenti studi, basati sull'analisi di alcune sequenze geniche di mtDNA, suggeriscono che, senza arrivare a parlare di sottospecie, vi fu sicuramente una suddivisione in tre (o forse quattro, ma il metodo non riesce ancora a chiarire quest'ipotesi) diversi grandi gruppi di popolazioni[13]. La reale esistenza dei gruppi sud-europeo (sud-iberico, subalpino, balcanico), centro-est europeo (dalla zona nord-iberica fino al mar Caspio) e medio-asiatico (fino ai confini orientali kazaki) in precedenza era stata frequentemente messa in discussione sulla base dei soli reperti fossili.
Tecnica, cultura e arte
L'uomo di Neanderthal inizia a evolvere in un contesto culturale acheuleano superiore, dove i manufatti bifacciali cambiano forma, migliorano la punta e diminuiscono di spessore.
Nell'industria litica compare la nuova tecnica di scheggiatura levalloisiana (da Levallois, alla periferia di Parigi). Da un nucleo litico iniziale, sgrossato fino a portarlo a una forma biconvessa, lateralmente su una faccia si staccano parallelamente a un piano di base schegge di forma regolare. Questa tecnica evolve e le forme chiamate amigdale (a mo' di mandorla) dell'Acheuleano scompaiono, anche se a sud del Sahara continuerà fino al 50 000 a.C. circa.
In Europa, territorio principale del Neanderthal, si parla di cultura musteriana, da ritrovamenti a Le Moustier, in Dordogna. Abbiamo punte triangolari, raschiatoi (per la preparazione delle pelli) molto rifiniti, dal bordo tagliente finemente ritoccato. Il Musteriano si articola in diverse culture, geografiche e cronologiche (Musteriano di tradizione acheuleana, Musteriano tipico, Musteriano denticolato, Charentiano, Musteriano pontiniano nel Lazio eccetera).
Pare accertato, con qualche residua incertezza, il passaggio successivo al Castelperroniano[14], con reperti affidabilmente attribuiti ai Neandertaliani come lamette litiche, manufatti in osso e ornamenti per il corpo.
Evoluzione culturale
Sull'evoluzione culturale di Homo neanderthalensis non vi è ancora una visione condivisa. A fronte di antropologi come Ian Tattersall, che non riconoscono il raggiungimento di livelli culturali che sconfinino dalla mera tecnologia e, in particolare, non condividendo il fatto che sia stata raggiunta la visione simbolica, intesa come dimensione simbolica nel senso psicologico, premessa a riti, arte, e comportamenti relativi, vi sono invece indizi di comportamenti culturali avanzati.

Le culture litiche che poi evolveranno (Castelperroniano, Aurignaziano e molto dubbiosamente Gravettiano, condivise sicuramente dai sapiens) sono quindi tuttora allo studio per la sicura eventuale attribuzione ai Neandertal; pare, con buona significatività, accertato il primo passaggio.
In sintesi ed a grandi linee, si può dire che la cultura neandertaliana dominante fu il Musteriano e che il limite convenzionale (attuale) superiore si situa fra il Castelperroniano e l'Aurignaziano. Molto diffuso era l'utilizzo delle pelli, anche per la costruzione di ripari estivi all'aperto, contrapponendosi alla pratica troglodita invernale. Si ritrovano strutture di pietre o di ossa atte ad assicurare i bordi delle pelli al suolo.
Abbondanti tracce di ocra rossa fanno pensare ad usi rituali e religiosi. Anche in tale ottica, si evidenzia l'inumazione come pratica diffusa, in fosse di forma ovale, con corredi funerari (cibo, corna e strumenti litici), spesso ricoperte da lastroni per sottrarre i corpi alle fiere, deposizioni di fiori (studi sui pollini in ritrovamenti in Asia Minore). Il fuoco, in cerchi di contenimento di pietre, è largamente utilizzato.
Secondo una ricerca in corso nel sito della grotta di Bruniquel in Francia, i neanderthalensis avrebbero realizzato, all'incirca 175 000 anni or sono, strutture complesse (la cui funzione è ancora oggetto di indagine) utilizzando stalagmiti appositamente spezzate e poi disposte in formazioni concentriche, dimostrando un'avanzata capacità organizzativa, costruttiva e, forse, simbolica[15].
Forse (gli studi sono ancora in corso), con i neanderthalensis si ha il primo esempio di strumento musicale non a percussione, ma intonato (in dettaglio, con quattro note compatibili[16], con la naturale scala diatonica greca), grazie al ritrovamento del cosiddetto flauto di Divje Babe (in Slovenia): un frammento di femore di orso delle caverne perforato regolarmente. Inizia anche l'arte figurativa in senso stretto, considerata prerogativa di sapiens, ma dalla stratigrafia recentemente attribuita anche ai neanderthalensis.
João Zilhão, professore all'Università di Lisbona, ha affermato che i Neanderthal avevano la capacità di usare simboli e pensare in modo astratto, basandosi sulle grotte in Spagna.[17]
I recenti progressi molecolari nello studio delle popolazioni neandertaliane e la loro localizzazione geografica, uniti a quelli sull'industria litica e degli altri manufatti, permetteranno in futuro di chiarire meglio i rapporti tra le diverse culture e la loro evoluzione nello spazio e nel tempo.
Nel complesso, la tecnologia dell'Homo neanderthalensis può riassumersi in:
- Asce a mano o amigdale: sono il resto di grossi noduli di selce, scheggiati ai bordi per ricavarne schegge più piccole.
- Punte di selce: da usare immanicate su pesanti bastoni usati come lance nella caccia a grossi animali.
- Denticolati, cioè schegge di selce senza punta col margine dentellato: sarebbero primitive seghe a mano, usate per lavorare legno, ossa e tendini.
- Raschiatoi: sono dei coltelli di selce da usare senza manico, per tagliare la carne.
- Flauti, cioè ossa lunghe forate, che sarebbero una sorta di accendini: nei fori venivano sfregati bastoncini di legno per accendere paglia e altro materiale infiammabile.
Un articolo pubblicato su Proceedings of the National Academy of Sciences il 13 novembre 2012 a cura di un gruppo del Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology di Lipsia[18], getta nuova luce sulla sempre più probabile attività simbolica e artistica dei Neandertal nel passaggio dal Paleolitico medio (MP) a Paleolitico superiore (UP), periodo caratterizzato dalla sostituzione degli ultimi Neandertal con gli esseri umani moderni in Europa tra 50 000 e 40 000 anni fa.
Le raccolte di manufatti del Castelperroniano (CP) trovati nella Francia centrale e nella Spagna settentrionale sono databili in questo intervallo di tempo. Fino ad ora, è il solo tipo di collezioni che ha consentito di associare i resti Neandertal con manufatti di stile UP. I manufatti CP includono anche ornamenti del corpo, praticamente sconosciuti in tutto il mondo Neandertal. Tuttavia, si è sostenuto che i manufatti CP non siano stati fabbricati da Neandertaliani, ma piuttosto i processi di formazione dei siti e la commistione degli strati abbiano portato all'associazione casuale di resti di Neandertaliani, assemblaggi CP, e gli ornamenti del corpo.
Fin dal Paleolitico medio, la pece di betulla fu largamente utilizzata dagli Uomini di Neanderthal come collante. Uno studio del 2019 ha dimostrato come queste prime produzioni di pece fossero realizzate attraverso una tecnica relativamente semplice, utilizzando dei focolari a cielo aperto[19].
Gli autori pubblicano una serie di datazioni al radiocarbonio ottenute con spettrometria di accelerazione di massa analizzando collagene osseo ultrafiltrato estratto da 40 frammenti di ossa ben conservate del tardo Musteriano, CP, e dagli strati Protoaurignaziani nel sito di Grotte du Renne (a Arcy-sur-Cure, Francia). I risultati sono incompatibili con l'ipotesi di commistione. Inoltre, si riporta una data direttamente calcolata su di un frammento di osso compatto di tibia di Neanderthal rinvenuto a Saint-Césaire (Francia). Questa data conferma l'assegnazione di manufatti CP agli ultimi Neanderthal dell'Europa occidentale. Inoltre, ed è ancora più importante, i dati stabiliscono che la produzione di ornamenti corporei nel CP è successiva all'arrivo degli umani moderni nelle regioni adiacenti dell'Europa. Questo nuovo comportamento potrebbe quindi essere il risultato di una diffusione culturale dai gruppi moderni ai gruppi di Neandertaliani.
Distribuzione territoriale
L'uomo di Neandertal è originario[20][21] dell'Europa occidentale e centrale, poi emigrato sulla via del Medio Oriente, sugli attuali territori di Iraq, Siria e d'Israele, con pochi individui fino in Asia centrale (Uzbekistan) e in Siberia.
L'evoluzione che ha condotto alla comparsa dell'Homo neanderthalensis, o «neandertalizzazione», è stata lenta e progressiva, da gruppi europei isolati (Homo erectus, Homo georgicus, Homo antecessor). Può essere seguita partendo dai pre-Neandertaliani e a seguire fino ai Neandertaliani recenti.
- I pre-Neandertaliani antichi
- La prima tappa corrisponde a fossili generalmente attribuiti a Homo heidelbergensis, possibile antenato, secondo alcune teorie anche dei sapiens moderni: è il caso dell'Uomo di Tautavel (−400 000 anni), rinvenuto in Francia, della mandibola di Mauer (−600 000 anni), trovato vicino a Heidelberg in Germania, o del cranio di Petralona (Grecia).
- I pre-Neandertaliani recenti
- La prima tappa corrisponde ai fossili di Swanscombe (Inghilterra), di Steinheim (Germania) o della Sima de los Huesos à Atapuerca (Spagna).
- I Neandertaliani antichi
- I successivi fossili con tratti innegabilmente Neandertaliani hanno un'età compresa tra −250 000 e −110 000 anni. Si può citare il cranio Biache-Saint-Vaast (Passo di Calais), i resti de la Chaise (Charente), la mandibole di Montmaurin (Alta-Garonna), i crani italiani di Saccopastore (Lazio) o l'abbondante materiale di Krapina in Croazia.
- I Neandertaliani classici
- I Neandertaliani tipici, con caratteri derivativi più marcati hanno un'età compresa tra −100 000 anni e −28 000 anni, data degli ultimi rinvenimenti fossili, e presumibilmente della loro sparizione.
Inoltre, oltre ai fossili di Neandertal stesso (circa −42 000 anni), si ricordano gli scheletri di La Chapelle-aux-Saints, di Moustier, di La Ferrassie,[22] di La Quina, di Saint-Césaire nel sud-ovest della Francia o della Spy in Belgio.
Gli ultimi Neandertaliani noti sono stati rinvenuti in Portogallo, in Spagna (Zafarraya −30 000 anni[23],[24]), in Croazia (Vindija, −32 000 anni[25],[26]) e nel nord-ovest del Caucaso (Mezmaiskaya, −29 000 anni). Come già visto queste date sono da considerare con precauzione, e sono spesso controverse.
Alcune ricerche condotte dal 1999 al 2005 sulla grotta di Gorham a Gibilterra suggeriscono che i Neandertaliani l'abbiano abitata dai −28 000 ai −24 000 anni[27],[28]. Dunque una lunga coabitazione con Homo sapiens, geograficamente presente dai −32 000 anni. Joao Zilhao, dell'università di Bristol critica fermamente questi ultimi risultati spostando all'indietro la data di estinzione a 37 000 anni fa. Le diverse teorie sulle date di estinzione e coabitazione con H.s.s. allo stato attuale non possono che definirsi in fase di sviluppo e verifica, ciò anche alla luce di sempre nuovi ritrovamenti, anche di terzi coabitatori cogeneri, nella fascia temporale dei 30 000 - 50 000 anni fa[29].
Antropologia fisica
Quella che segue è una lista di tratti fisici che distinguono i Neandertal dagli esseri umani moderni; comunque, non tutti possono essere utilizzati per distinguere specifiche popolazioni di Neandertal, provenienti da varie aree geografiche o periodi di evoluzione, da altre specie di uomini estinte. Inoltre, molti di questi tratti si manifestano occasionalmente negli uomini moderni, specialmente tra certi gruppi etnici. Non si sa ancora nulla riguardo al colore della pelle, o alla forma di parti morbide come gli occhi, le orecchie e le labbra dei Neandertal, se non per modellistica.
Comparati ai moderni esseri umani i Neandertal erano di costituzione più robusta e avevano caratteri morfologici distintivi, specie per quanto riguarda il cranio, che gradualmente accumulava più aspetti derivati (in senso cladistico), specialmente in regioni geografiche relativamente isolate. Anche qui è da notare che le popolazioni anatomicamente moderne del paleolitico medio (vicino oriente come a Skul e Qafzeh), erano ugualmente massicce. Si pensa che la loro statura relativamente alta ma robusta sia un adattamento al clima freddo dell'Europa durante il Pleistocene, (legge di Allen) anche se in realtà questi tratti paiono già presenti in periodi caldi.
La seguente tabella è una sintesi, con integrazioni, della review su Sc. American di E. Trinkaus & W.W. Howells sul Neandertal.
| Cranico | Post-cranico |
|---|---|
| Fossa soprainiaca, una scanalatura sopra l'inion o linea superiore della nuca. | In genere sensibilmente robusto, ma sapiens coevi erano similmente robusti. |
| Bozzo occipitale, una protuberanza dell'osso occipitale. | Estremità delle dita (tuberosità) larghe e arrotondate, inserzioni robuste dei relativi tendini. |
| Prognatismo, margine del I molare più avanzato dello zigomo | Gabbia toracica tendenzialmente cilindrica |
| Teschio basso piatto (platicefalia) ed elongato antero posteriormente | Rotule grandi |
| Base cranica piatta, vista posteriormente di forma tondeggiante, contrapposta a quella sapiens pentagonale | Ossa del collo lunghe |
| Toro sopraorbitale, un'arcata sopraciliare prominente, trabecolata (spongiforme) | Scapola con cavità dorsale, contrapposta a quella moderna con cavità in prevalenza (4/5) ventrale, per una migliore inserzione del muscolo piccolo rotondo |
| 1200–1750 cm³ di capacità cranica (10% maggiore della media umana attuale) | Femore spesso e incurvato, indicante una buona potenza muscolare |
| Mento osseo poco prominente, in realtà spesso mascherato dal prognatismo; negli esemplari più recenti il carattere non si evidenzia | Tibia e fibula corte, idem radio e ulna |
| Cresta del processo mastoideo dietro l'apertura auricolare | Osso pubico di entrambi i sessi gracile e allungato (ramo pubico superiore), probabile aumento dimensionale del canale del parto |
| Nessuna scanalatura sui denti canini | Espansione delle superfici articolari dell'astragalo |
| Diastema posteriore al terzo molare, conseguenza dell'allungamento mascellare | Robuste inserzioni muscolari sull'arcata plantare del piede (forte spinta nella corsa) |
| Proiezioni ossee ai lati dell'apertura nasale | |
| Particolare forma del labirinto, nell'orecchio interno | |
| Grande forame mandibolare per il circolo ematico fornito di ponte osseo, nei sapiens in genere di doccia. | |
| Naso ampio ma non schiacciato |
Craniometria
Un'accurata statistica, basata sull'analisi multivariata di forma e dimensioni del cranio, tuttavia, pur rilevando un habitus tipico del Neandertal, mostra sorprendenti vicinanze con crani attuali di forme estreme (C.Stringer, del British Museum).
Pigmentazione
Da uno studio del 2001 alcuni commentatori speculano sul fatto che i Neandertal dimostrino un tipo a capelli rossi, condividendone l'eredità con uomini attuali di tipo lentigginoso e di pelo rosso;[30] in ogni caso altri ricercatori dell'epoca dissentivano.[31] Gli studi molecolari del 2007 citati nel paragrafo iniziale Sintesi dell'aspetto esteriore sciolgono comunque ogni dubbio, ovvero sulla base dello studio sulle varianti dei melanocortin 1 receptor (mc1r), (o melanocyte-stimulating hormone receptor (MSHR), melanin-activating peptide receptor, o melanotropin receptor) una classe di recettori cellulari, recettori accoppiati a proteine G, che sono ligandi di ormoni peptidici Ipofisari, si deduce che la ridotta funzionalità di questi porti a sviluppare un fenotipo poco pigmentato a pelle chiara e capelli rossi. Lo studio è stato condotto, con tecniche di amplificazione genica su un gene responsabile della pigmentazione umana, appunto quello relativo al melanocortin 1 receptor (mc1r), che codifica sette proteine eterometriche transmembrana GTP-ligande (G protein)–coupled receptor (GPCR). I capelli rossi e la pelle chiara risultano da alleli che comportino complete o parziali perdite di funzionalità dell'MC1R umano (huMC1R), alterando il bilancio nella sintesi eumelanina-feomelanina. I campioni provenivano da DNA ben conservato (sulla base di dati relativi ad abbondante contenuto amminoacidico e alla sua scarsa racemizzazione) di un esemplare dei Monti Lessini (in Italia) e dell'esemplare El Sidrón 1252 (Cueva de El Sidrón, in Spagna nelle Asturie).
Sviluppo fisico
Un'altra differenza secondo i paleoantropologi, potrebbe essere stata nella crescita: analizzando la crescita del primo molare comparata con quello della dentatura di Homo sapiens, si ritiene che i Neandertal crescessero più velocemente di quanto facciano gli uomini moderni, raggiungendo l'età adulta già a 15 anni. Questa crescita rapida, stando a quanto riportano gli esperti, è una caratteristica tipica dei primi ominidi[non chiaro]. Tempi di sviluppo più rapidi, secondo gli studiosi, sembrano essere stati una necessità per una popolazione che viveva in terre molto fredde e inospitali, dove la mortalità infantile era molto elevata e l'aspettativa di vita breve.
Ibridazione con Homo sapiens
|
Questa voce o sezione sull'argomento antropologia è ritenuta da controllare.
|

Nel corso della seconda metà del XX secolo si è assai dibattuto in ambito accademico se l'uomo di Neandertal sia da considerare una sottospecie estinta di Homo sapiens (denominata Homo sapiens neanderthalensis), con la conseguente possibilità di incrocio e ibridazione tra i due, oppure se sia una specie autonoma (Homo neanderthalensis). Comunque sia, i due gruppi convissero per migliaia di anni in Europa e Asia Minore. Indagini genetiche eseguite nel 2008 su DNA fossile recuperato su alcuni resti di uomo di Neandertal, sembravano indicare che i due tipi umani parzialmente contemporanei appartenessero a due diverse specie,[32] tuttavia più recenti scoperte riguardanti il genoma umano sembrerebbero in grado di accertare che neanderthalensis e sapiens apparterrebbero alla stessa specie e che le differenze anatomiche e somatiche sarebbero dovute a cause climatiche, ambientali, morfologiche e di alimentazione[33].
Analisi del DNA
Il biologo molecolare svedese Svante Pääbo ha testato almeno 70 campioni provenienti da Neandertaliani prima di trovarne uno sufficiente per le analisi del DNA; inizialmente sequenze da ossa di 38 000 anni provenienti dal sito croato di Vindija mostrano una condivisione del 99,5% del DNA con sapiens: i due appartenenti al genere Homo da ciò divergerebbero da circa 500 000 anni. Un articolo sulla rivista Nature stima la divergenza a 516 000 anni, dove le prove paleontologiche, sufficientemente concordi, mostrano un periodo di 400 000 anni. Svante Pääbo ha proposto un progetto per la ricostruzione totale del patrimonio genetico di Homo neanderthalensis. Edward Rubin del Lawrence Berkeley National Laboratory di Berkeley, California dimostra che il genoma dei Neandertal è simile al nostro dal 99,5 al 99,9%.[34][35]
Nel novembre 2006, un nuovo articolo viene pubblicato sulla rivista scientifica Proceedings of the National Academy of Sciences, dove un gruppo di ricerca europeo ripropone il tema dell'ibridazione[36]. Contemporaneamente il 16 novembre 2006 Science Daily pubblica la tesi opposta, elaborata dal Berkeley Lab e dal Joint Genome Institute, secondo cui non vi sarebbero evidenze di incroci tra i due gruppi umani e la sovrapposizione genetica non supererebbe il 99,5%.
Edward Rubin, direttore di entrambi gli istituti, conclude:
| (EN)
«While unable to definitively conclude that interbreeding between the two species of humans did not occur, analysis of the nuclear DNA from the Neanderthal suggests the low likelihood of it having occurred at any appreciable level.» |
(IT)
«Anche se non siamo in grado di affermare definitivamente che l'ibridazione tra le due specie non abbia avuto luogo, l'analisi del DNA nucleare del Neandertal suggerisce una bassa probabilità che questo sia avvenuto a livello apprezzabile.» |
La più accurata analisi molecolare condivisa a livello internazionale e attualmente confermata, stima che l'H. sapiens e l'H. neanderthalensis si separarono non più di 800 000 anni fa e non vi sono ancora prove evidenti accettate dalla maggioranza della comunità scientifica su un eventuale ricongiungimento.[37] Attualmente la comunità scientifica, sulla base di alcuni studi di Svante Pääbo dell'Istituto Max Planck di Lipsia, considera comunque possibile tra le varie ipotesi, un ricongiungimento genetico tra le due specie, sulla base della presenza, nel genoma di parte della popolazione umana attuale (esclusi gli africani), di circa l'1-4% di materiale genetico compatibile con quello neandertaliano.
In un ulteriore articolo di Svante Pääbo, in collaborazione con il Dipartimento di Genetica di Harvard e il MIT, viene proposta una datazione dell'ultimo periodo di scambio di geni tra Neandertal e umani moderni (H. sapiens sapiens)[38].
Una delle scoperte chiave dell'analisi del genoma di Neandertal è che i Neandertal condividono più varianti genetiche con non-africani che con gli africani. Questa osservazione è in linea con due ipotesi: 1) l'incrocio tra Neandertal e gli esseri umani moderni è avvenuto dopo che gli umani moderni sono usciti dell'Africa o 2) la struttura delle popolazioni degli antenati di Neandertal e di quelli degli esseri umani moderni erano in parte simili. Queste ipotesi possono fare previsioni diverse sulla data dell'ultimo scambio di geni tra gli antenati di Neandertal e gli umani moderni non africani. Gli autori stimano questa data misurando il grado di linkage disequilibrium (LD) (spesso definita "associazione allelica"; particolari alleli in due o più loci vicini mostrano associazione allelica se si verificano con frequenze significativamente diverse da quelle previste in base alle frequenze alleliche dei singoli) tra il genoma degli attuali europei e trovano che l'ultimo flusso genico da Neandertal agli europei si è probabilmente verificato 37 000-86 000 anni fa (BP), e più probabilmente 47 000-65 000 anni BP. Ciò supporta l'ipotesi di un incrocio recente e suggerisce che l'incrocio si è verificato quando gli esseri umani moderni portatori delle tecnologie del Paleolitico superiore incontrarono i Neandertal a seguito della loro espansione fuori dall'Africa.
Le tracce neandertaliane più recenti
Si dibatte lungamente sugli ultimi rappresentanti noti dei nostri parenti più prossimi. Quest'argomento, assieme al mistero della scomparsa della popolazione neanderthaliana, è forse uno dei più controversi della scienza paleoantropologica e il più soggetto a evoluzione. Le datazioni tramite spettrometria di massa da parte di Fred Smith ed Erik Trinkaus, (Northern Illinois University e Washington University rispettivamente), da scavi di Vindija (Croazia) portano a 28 000 anni i reperti più recenti, con articoli pubblicati a cavallo del 2000. Nuove datazioni ricollocano indietro[39] a 32 000 anni i reperti. Vengono scoperti altri fossili recenti sulle coste atlantiche del Portogallo, e si riscontrano evoluzioni morfologiche verso una maggiore modernità. Contemporaneamente viene messa in discussione l'associazione di alcune culture litiche (Aurignaziano, Musteriano) ai vari Homo.
La scomparsa dell'intera popolazione neanderthaliana

Si dibatte sulla completa scomparsa della popolazione, non perfettamente spiegabile sulla base delle sole caratteristiche fisiche degli individui. Si trattava di una specie lungamente adattata all'ambiente colonizzato, con un volume cranico pari o superiore ai sapiens attuali, e di cultura tecnica almeno inizialmente sovrapponibile nelle due popolazioni. Le prime ipotesi teorizzate parlano di lenta ibridazione con Sapiens moderni, eliminazione fisica (genocidio), competizione, o selezione sessuale. La difficoltà ad analizzare lo scarso materiale genetico sopravvissuto completa il quadro[40]. Certamente la lunga coesistenza di uomo di Neandertal e uomo moderno pongono sul tavolo della discussione molti problemi irrisolti e l'argomento è in costante riscrittura.
La teoria del "rimpiazzamento"
Nel 2005 sul Journal of Economic Behaviour and Organization Jason Shogren, economista dell'Università del Wyoming di Laramie, pubblica con i suoi collaboratori un articolo in cui avanza una teoria sulla scomparsa dell'uomo di Neandertal[41]. Lo studioso avanza l'ipotesi che H. neanderthalensis si sia dovuto scontrare con la particolare cultura dell'H. sapiens: questa cultura si basava su tecniche avanzate di commercio, cosa che portava più tempo libero rispetto a una cultura basata sulla caccia. Il tempo libero ottenuto avrebbe permesso lo sviluppo di specializzazioni non strettamente legate alla sussistenza, come costruire utensili sempre più complessi o dedicarsi all'arte. La complessità e la versatilità di una tale cultura avrebbe avuto esito fatale per la più "tradizionale" cultura dei Neandertal.
Stephen Kuhn e Mary Stiner dell'università dell'Arizona, sulla rivista Current Anthropology, propongono, documentandola, la tesi per cui la principale causa di estinzione fu la mancata suddivisione dei lavori tra i sessi. I più organizzati sapiens, più efficientemente, poterono competere affidando alle donne compiti stanziali, e meno gravosi, affidando ai maschi i ruoli di cacciatori e approvvigionatori di materiali. La prole, protetta e anch'essa stanziale, avrebbe avuto più possibilità di sopravvivenza[42]. Un'altra ipotesi sulla loro estinzione, fa riferimento invece ai drastici cambiamenti nella composizione della magnetosfera terrestre, dovuti all'azione di forti variazioni del campo magnetico terrestre, con conseguente variazione della radiazione solare che avrebbe comportato, a sua volta, mutazioni negative per la specie, fino a portarla all'estinzione.[43]
Due caratteristiche dei neanderthalensis vanno rimarcate. La prima è che il loro fisico era strutturato per esprimere al meglio la forza, mentre quello di alcuni, ma non tutti, i sapiens loro contemporanei come i Cromagnonoidi, a gambe più lunghe e a bacino più stretto e compatto, privilegiava le capacità di resistenza nella corsa. Quindi quei sapiens sarebbero stati più resistenti nel percorso di sensibili distanze con quel tipo di andatura, ma svantaggiati nella forza fisica e nella lotta. Una caratteristica che si pensava differenziasse sapiens e Neandertal, la diversa alimentazione, con sapiens marcatamente onnivori e Neandertal carnivori si è variamente rivelata parziale e dipendente esclusivamente da singole situazioni. Entrambi ad esempio erano specie ben adattate agli ambienti costieri[44] con un'alimentazione basata su frutta e verdura, prodotti della pesca, raccolta di molluschi e caccia[45]. Soggetta a periodiche discussioni è anche la capacità dei Neandertal di cacciare grosse prede (come la megafauna); mentre esistono prove che anche i più antichi esponenti della nostra specie utilizzassero zagaglie e altri ordigni da lancio, non si sa se altrettanto facessero i Neandertal, i quali avrebbero pertanto dovuto aggredire le prede (e eventuali nemici) da distanza ravvicinata, esponendosi a maggiori rischi per la propria incolumità. I Neandertal, almeno alcune popolazioni, erano comunque prettamente carnivori[46] e superpredatori.
La teoria dell'"ibridazione"
Uno studio pubblicato su Science Advance nel 2020 afferma che circa 50.000 anni fa, mentre Homo sapiens si diffondeva fuori dell'Africa, avvennero accoppiamenti fertili con popolazioni neandertaliane e denisoviane sino al loro assorbimento e sostituzione con popolazioni tipicamente sapiens ma nel cui genoma rimase traccia dell'avvenuta mescolanza genetica.[47]
Nel 2010 sono stati pubblicati i risultati di una ricerca quadriennale che mirava a mappare il genoma dei Neanderthal, è stato comparato del DNA raccolto da dei fossili di Neanderthal con del Dna degli umani contemporanei, e i risultati hanno stupefatto la comunità scientifica, nel DNA unico condiviso delle moderne popolazioni una porzione rilevante tra 1 e il 4% era costituito da DNA Neanderthaliano.[48]
Ricostruzioni dell'aspetto, secondo la scienza e gli stereotipi


|
«Se [un uomo di Neandertal] si potesse reincarnare e porre nella metropolitana di New York, opportunamente lavato, sbarbato e modernamente vestito, si dubita che potrebbe attirare più attenzione di alcuni dei suoi altri abitanti.» |
| (William Straus[49]) |

Molto ma non tutto si può dire del vero aspetto di questo ominide ricostruendolo dai suoi resti fossili. La scienza contemporanea ha messo a punto varie tecniche per la ricostruzione delle parti non fossilizzabili, i tessuti molli, a partire da elementi quali le inserzioni tendinee sulle ossa, le linee di forza sulle stesse, i livelli di consunzione dei denti e altro ancora. La biologia molecolare ha poi fornito ulteriori elementi per valutare l'espressione genica di caratteri non conservabili, appunto come il colore dei capelli. Il Neandertal possedeva la tecnologia necessaria a confezionare indumenti; a tale scopo utilizzava prevalentemente pelli, per la concia delle quali costruiva utensili quali i raschiatoi musteriani. Le zone climatiche frequentate imponevano sicuramente l'uso di coperture, e possiamo rappresentare il Neandertal tipico prevalentemente vestito.
Oltre a ciò si segnala la capacità simbolica e artistica, che ha portato all'uso, almeno episodico, di monili e pendagli[50]. In passato, al contrario, è spesso stato rappresentato come lo stereotipo dell'uomo cavernicolo, tale ricostruzione ripete né più né meno la figura dell'uomo selvatico presente nella tradizione popolare europea e raffigurata sin dal Medioevo. Secondo alcune interpretazioni di studiosi di criptozoologia, non di antropologi o paleontologi, poteva però essere di aspetto scimmiesco[51], dotato di un fitto villo pilifero distribuito su tutta la superficie del corpo che lo proteggeva dal clima freddo europeo.
Un'immagine da alcuni supposta e interpretata di Neandertal è una rappresentazione preistorica nella caverna di Isturits sui bassi Pirenei francesi, frequentata da 80 000 a 10 000 anni fa prima dai Neandertal e poi dai Cro-magnon. Vi compare la rappresentazione di una testa-trofeo di un essere antropomorfo con testa ovale senza fronte e mento, collo grosso, occhi a mandorla infossati sotto grandi arcate frontali, con un grosso naso, orecchie piccole, con capelli corti a spazzola e dritti a cresta sulla testa che proseguivano sulla schiena in una sorta di criniera equina, e peli fitti simili su quasi tutta la faccia e collo simili al manto di un cervo.[senza fonte]
Storia dei principali reperti fossili europei

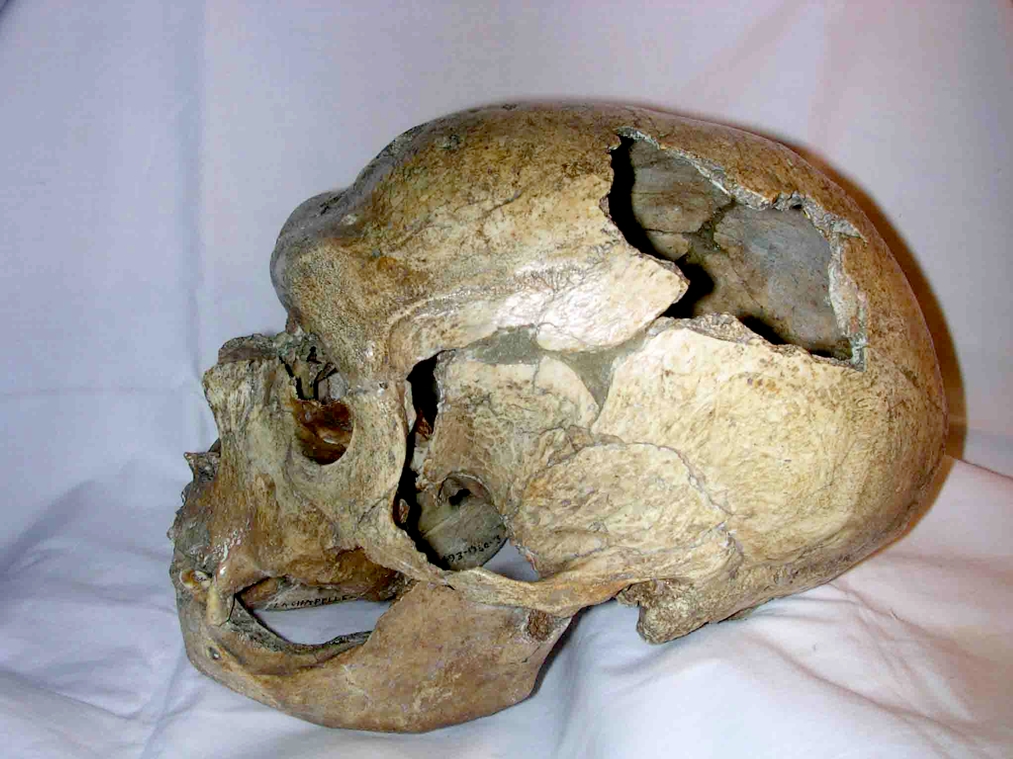
I resti rinvenuti nel 1856 da Johann Fuhlrott nella valle di Neander consistevano nella parte superiore del cranio, alcune ossa, parte dell'osso pelvico, alcune costole, e ossa del braccio e della spalla.
In precedenza erano stati scoperti altri fossili, infatti già nel 1829 nel Belgio venne trovato parte di un cranio di un bambino di due anni e mezzo. Questi, però, venne riconosciuto come arcaico soltanto nel 1836. Nel 1848 a Gibilterra venne trovato un cranio adulto, poi battezzato Gibraltar 1, ma la sua esistenza rimase sconosciuta alla scienza fino al 1864, quando venne riconosciuto come appartenente a un individuo Neandertal.
Altri due scheletri di Homo neanderthalensis, di almeno 60 000 anni, vennero trovati in Belgio nel 1886 da Marcel de Puydt e Max Lohest. Altri rinvenimenti importanti vennero fatti in Croazia nel 1899 da Dragutin Gorjanovic-Kramberger e nel 1908 in Francia a La Chapelle-aux-Saints da Jean Bouyssonie che rinvenne lo scheletro di un uomo anziano, risalente a 50 000 anni, il cui volume cranico era di ben 1620 centimetri cubi.

Nel 1939 venne rinvenuto nella grotta Guattari a San Felice Circeo, un cranio presumibilmente appartenente a H. neanderthalensis.
Nel secondo dopo guerra emersero ancora altri resti importanti; tra il 1953 e il 1960 nella grotta di Shanidar in Iraq vennero scoperti 9 scheletri di uomini di Neandertal, risalenti a un periodo compreso tra i 70 e i 40 000 anni fa, e nel 1979 nel villaggio di Saint-Césaire in Francia uno scheletro completo risalente a 35 000 anni fa.
Nel 1868 a Cro-Magnon in Francia vennero trovati da alcuni operai i resti di un uomo risalenti a 28 000 anni fa; era venuto alla luce uno dei più antichi progenitori della nostra specie (Homo sapiens). Era un rappresentante della nostra specie umana che, proveniente dall'Africa o dall'Asia, migrando stava insediandosi in Europa, confinando verso la penisola iberica gli ultimi Neandertal.
Resti neandertaliani rinvenuti in territorio italiano
I resti portati alla luce in Italia non sono molto numerosi rispetto all'Europa continentale. Si distingue in ogni caso tra ritrovamenti fossili (rari) e ritrovamenti di tracce e manufatti, non sempre associati. Nel secondo caso, alcuni siti sono discussi; alcune culture, infatti, furono condivise con soggetti appartenenti al genere Homo. Per i siti composti da soli reperti litici si veda la voce Paleolitico.
- Si riportano i siti tardo pleistocenici con reperti ominidi fossili in Italia, definibili di caratterizzazione Neandertaliana
Piemonte
- Monte Fenera (Ciota Ciara, Ciutarun) Vercelli: frammenti cranici, denti
Liguria
- Grotte dei Balzi Rossi. Caverne e grotte a strapiombo sul mare, rosse dalla ricchezza di composti ferrosi nel terreno, vicino al confine francese, monte Bellinda a Grimaldi, frazione di Ventimiglia. Ritrovamenti litici appartenenti al Musteriano. Abitate in successione da diverse culture e popolazioni, dal paleolitico medio fino al neolitico. Sepolture, principalmente Sapiens, utensili litici, arte rupestre, spesso di incerta attribuzione. I pochi fossili ritrovati sono di tipo pre-neandertaliano o neandertaliano antico medio pleistocenico.
- Caverna delle Fate, Finale Ligure (Savona)[52] manufatti Litici di tipo Levalloisiano e Musteriano datazione 100 000 - 50 000 a.C., ancora allo studio per una datazione palinologica. Frammenti ossei vari, di scavo anni novanta; si son conservati resti ossei, almeno di cinque Neandertal bambini e adulti, caso unico nell'archeologia dell'Italia settentrionale[53]. Nella vicina Arma delle Mànie si ritrovano ugualmente tracce neandertaliane.
- Grotte di Toirano, tra Albenga e Savona, molto vaste e concrezionate. Nella Grotta delle streghe, o della Basura, si rinviene un'impronta fossile di un piede giovane[54], (ma alcune datazioni radiometriche correlate alle impronte hanno stimato un'età di 12 000 anni, nel qual caso sarebbero allo stato attuale delle conoscenze, attribuibili solamente a Homo sapiens sapiens)[senza fonte] e molti resti animali (Ursus spelaeus). Le grotte e le loro testimonianze fossili, frequentate sicuramente dall'Homo heidelbergensis sino ai sapiens del neolitico, sono tuttora allo studio.
Veneto
Toscana
Lazio
- Monte Circeo, Grotta Guattari, (Latina). Il ritrovamento il 24 febbraio 1939 da parte del prof. Alberto Carlo Blanc fece a lungo discutere per l'interpretazione fortemente discussa di un cranio con segni di aggressione e svuotamento encefalico. Per mezzo secolo si discusse sull'antropofagia rituale della specie. Studi tafonomici successivi spiegarono lo svuotamento encefalico come azione di iene. Scavi archeologici tra il 2019 e il 2021 hanno riportato alla luce resti di altri 9 individui, portando ad 11 il numero di Homo neanderthalensis ritrovati nel sito.[59]
- Monte Circeo, (Latina), Grotta del Fossellone: mandibola.
- Monte Circeo, (Latina), Grotta Breuil: al suo interno, gli scavi del 1986 hanno rinvenuto una porzione postero-inferiore di parietale, una corona di primo molare inferiore di adulto e il terzo molare inferiore di un individuo giovane (circa 13 anni)[senza fonte].
- Roma, Cava di Saccopastore. Ritrovamento nel 1929 e 1935 di due soggetti, di tipo arcaico databili circa 120 000 anni[60].
Abruzzo
Campania
- Grotta Taddeo, Marina di Camerota, (Salerno): denti.
- Il Molare, a Scario, dove si ritrova una mandibola.
- Grotta di Vallicelli, a Monte San Giacomo, industria litica.
Puglia
- Sulla costa del Salento (Grotta Romanelli).
- A Capo di Leuca, (Lecce), la Grotta delle Tre Porte. Nella sala denominata 'Antro del bambino' insieme a resti di fauna pleistocenica si ritrova il molare fossile appartenuto a un bimbo Neandertal di circa dieci anni[61][62][63].
- Grotta di Santa Croce, Bisceglie, (Bari): femore e frammento cranico.
- Grotta di Lamalunga, nei pressi di Altamura (Bari): scheletro completo, denominato Uomo di Altamura).
- Grotta del Cavallo, Nardò, (Lecce)
- Grotta-Riparo Uluzzo, Nardò, (Lecce): denti.
- Fondo Cattìe, Maglie, (Lecce)[64],[65].
- Cava Nuzzo, Melpignano (Lecce): dente.
Calabria
- Contrada Ianni di San Calogero, Nicotera, (Vibo Valentia): porzione cranica.
- San Francesco di Archi, (Reggio Calabria): mandibola.
Solamente reperti litici
Lombardia
- Grotta dell'Orso (Monte Generoso – San Fedele Intelvi – CO), piccoli reperti in selce.[66]
- Cascina Barbabella di Bagaggera (Parco regionale di Montevecchia e della Valle del Curone – Rovagnate – LC), resti di focolari e concentrazioni di manufatti in selce[senza fonte]
- Buco del piombo[67] (Erba – CO)
Toscana
- Cava di La Pietra, Roccastrada (provincia di Grosseto)[68].
Nella cultura di massa
- Nella serie a fumetti Martin Mystère, creata da Alfredo Castelli ed edita dalla Bonelli, Java, il compagno di avventure del protagonista, è un uomo di Neandertal appartenente a una tribù della Mongolia, unica sopravvissuta all'estinzione della specie. Le caratteristiche fisiche sono leggermente differenti dalla realtà, in particolare è più alto e molto prestante fisicamente, mentre permane la caratteristica del mutismo, infatti Java si esprime con suoni gutturali e a gesti, aiutandosi anche con la lingua dei segni.
- In Far Cry Primal, gli Udam sono descritti come neandertaliani, benché questi fossero già estinti.
- In Topolino all'età della pietra (The Land of Long Ago), riduzione a fumetti del romanzo Il mondo perduto di Arthur Conan Doyle, il professor Ossivecchi ha come assistente Oompa, un uomo di Neanderthal, che si esprime a gesti e al quale il professore ha insegnato qualche parola.[69]
Nella letteratura e nel cinema
Numerosi romanzi e racconti vedono tra i protagonisti uno o più Neandertaliani:
- Jean M. Auel, Ayla figlia della Terra.
- L. Sprague de Camp, L'uomo nodoso (The gnarly man), 1939.
- Lester del Rey, Il giorno è compiuto (The Day Is Done), 1939.
- William Golding, Uomini nudi (The inheritors), 1955.
- Michael Crichton, Mangiatori di morte (Eaters of the Dead), 1976.
- Robert J. Sawyer, Trilogia dei Neanderthal (Neanderthal Parallax), 2002-2003.
- Isaac Asimov e Robert Silverberg, Il figlio del tempo (The Ugly Little Boy), 1992, basato sull'omonimo racconto di Asimov del 1958
- Stanis Mulas, L'uomo dei Balzi Rossi, 1988.
- Ben Bova, Orion, 1984.
Nel cinema:
- La guerra del fuoco (La guerre du feu) di Jean-Jacques Annaud (1981)
- Il 13º guerriero di John McTiernan (1999), adattamento cinematografico di Mangiatori di morte di Crichton
- L'era glaciale di Chris Wedge (2002)
- Una notte al museo di Shawn Levy (2006)
- Una notte al museo 2 - La fuga di Shawn Levy (2009)
- Ao, l'ultimo dei Neanderthal di Jacques Malaterre (2010)
- I Croods di Kirk DeMicco e Chris Sanders (2013)
- Una notte al museo 3 - Il segreto del faraone di Shawn Levy (2014)
- Il viaggio di Arlo di Peter Sohn (2015)
Note
- Il termine Neanderthal, scritto con "h", è corretto solo nel nome latino Homo neanderthalensis in quanto, secondo l'ortografia tedesca attuale, la parola Tal (valle) ha perso la "h" che aveva prima delle regolamentazioni linguistiche sancite dalla Conferenza ortografica del 1901(DE) Thal - Wiktionary, su de.wiktionary.org. URL consultato il 14 marzo 2017.«Thal ist eine alte Schreibweise von Tal. Sie ist gemäß den Beschlüssen der Orthographischen Konferenz von 1901 seither nicht mehr korrekt.»
- Neanderthal Museum, Storia del ritrovamento
- Richard E. Green et al, A Draft Sequence of the Neanderthal Genome, in Science, vol. 328, n. 5979, pp. 710–722, DOI:10.1126/science.1188021.
- Paul Rincon, Neanderthal genes 'survive in us', su BBC News, BBC, 6 maggio 2010. URL consultato il 7 maggio 2010.
- Le ossa lunghe sono quelle che si sviluppano in lunghezza e sono, in particolare:
- femore, osso della gamba;
- tibia, osso della gamba;
- perone, osso della gamba;
- radio, osso del braccio;
- ulna, osso del braccio;
- omero, osso del braccio
- H. Helmuth, Body height, body mass and surface area of the Neanderthals., in Z Morphol Anthropol, vol. 82, n. 1, 1998, pp. 1-12, PMID 9850627.
- Froehle, Andrew W; Churchill, Steven E (2009), Energetic Competition Between Neandertals and Anatomically Modern Humans, (PDF), PaleoAnthropology: 96–116, 2009; collegamento consultato il 31 ottobre 2011.
- (EN) M.S. Ponce de León, L. Golovanova, V. Doronichev, G. Romanova, T. Akazawa, O. Kondo, H. Ishida e C.P. Zollikofer, Neanderthal brain size at birth provides insights into the evolution of human life history, in PNAS, n. 105, 2008, pp. 13764-13768.
- (EN) Carles Lalueza-Fox et al., A Melanocortin 1 Receptor Allele Suggests Varying Pigmentation Among Neanderthals., su sciencemag.org, 25 ottobre 2007.
- Articolo di sintesi [collegamento interrotto], su repubblica.it.
- (EN) Laurent Excoffier, Neanderthal Genetic Diversity: A Fresh Look from Old Samples Current Biology, vol. 16, 22 agosto 2006, pp. R650-R652.
- (EN) Carles Lalueza-Fox, In Neanderthals, there was probably the whole range of hair colour we see today in modern European populations, from dark to blond right through to red.
- (EN) V. Fabre, S. Condemi e A. Degioanni, Genetic Evidence of Geographical Groups among Neanderthals, in PLoS ONE, 4(4), 2009, p. e5151, DOI:10.1371/journal.pone.0005151.
- (EN) Jean-Jacques Hublina, Sahra Talamo, Michèle Julien, Francine David, Nelly Connetb, Pierre Bodub, Bernard Vandermeerschd e Michael P. Richardsa, Radiocarbon dates from the Grotte du Renne and Saint-Césaire support a Neanderthal origin for the Châtelperronian, in PNAS, vol. 109, n. 46, 29 ottobre 2012, pp. 18743-18748, DOI:10.1073/pnas.1212924109.
- (EN) Jacques Jaubert, Sophie Verheyden e Dominique Genty, Early Neanderthal constructions deep in Bruniquel Cave in southwestern France, in Nature, advance online publication, 25 maggio 2016, DOI:10.1038/nature18291. URL consultato il 26 maggio 2016.
- Bob Fink, Neanderthal Flute, Greenwich, 1997, ISBN 0-912424-12-5. WorldCat.org)
- L'Origine dell'Arte e i Primi Esempi di Arte Paleolitica, su ulukayin.org, 2 novembre 2022. URL consultato il 13 novembre 2022.
- J.J. Hublin et al., Radiocarbon dates from the Grotte du Renne and Saint-Cesaire support a Neanderthal origin for the Chatelperronian, in Proc Natl Acad Sci U S A., vol. 109, n. 46, 13 novembre 2012, pp. 18743-8, DOI:10.1073/pnas.1212924109.
- (EN) Schmidt, P., Blessing, M., Rageot, M., Iovita, R., Pfleging, J., Nickel, K. G.; Righetti, L. & Tennie, C., Birch tar extraction does not prove Neanderthal behavioral complexity, in PNAS, 19 agosto 2019, DOI:10.1073/pnas.1911137116.
- (FR) J.-J. Hublin, Y. Coppens e P. Picq, Origine et évolution des Néandertaliens, in Aux origines de l'humanité, vol. 1, Fayard, 2001.
- (FR) J. Jaubert, Chasseurs et artisans du Moustérien, in La Maison des Roches, 1999..
- Anche i Neandertal seppellivano i loro morti, su lescienze.it, 9 dicembre 2020. URL consultato il dicembre 2020.
- (EN) J.J. Hublin, C. Barroso Ruiz, P. Medina Lara, M. Fontugne e J.-L. Reyss, The Mousterian site of Zafarraya (Andalucia, Spain): dating and implications on the palaeolithic peopling processes of Western Europe, in Comptes rendus de l'Académie des sciences, 321 (IIa), 1995, pp. 931-937.
- (EN) J.-J Hublin e E. Trinkaus, The Mousterian human remains from Zafarraya (Andalucia, Spain), in American Journal of Physical Anthropology, supplemento 26, 1998, pp. 122-123.
- (EN) F.H. Smith, E. Trinkaus, P.B. Pettitt, I. Karavanić e M. Paunovic, Direct radiocarbon dates for Vindija G1 and Velika Pećina Late Pleistocene hominid remains, in PNAS, vol. 96, n. 22, 26 ottobre 1999, pp. 12281-12286.
- T. Higham, C.B. Ramsey, I. Karavanić, F.H. Smith e E. Trinkaus, Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neanderthals, in PNAS, vol. 103, n. 3, 17 gennaio 2006, pp. 553-557.
- (EN) C. Finlayson et al., Late survival of Neanderthals at the southernmost extreme of Europe, in Nature, 13 settembre 2006.
- (EN) D. Brill, Neanderthal's last stand, in news@nature.com, 13 settembre 2006.
- Johannes Krause et al. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia, Nature advance online publication 24 March 2010 | doi:10.1038/nature08976
- Red-Heads and Neanderthals, su gktgazette.com, maggio 2001. URL consultato il 28 ottobre 2005 (archiviato dall'url originale il 18 ottobre 2005).
- Nicole's hair secrets, su arts.telegraph.co.uk, 10 febbraio 2002. URL consultato il 2 novembre 2005 (archiviato dall'url originale il 24 febbraio 2006).
- Roxanne Khamsi, Neanderthal DNA illuminates split with humans, su newscientist.com, 11 ottobre 2006. URL consultato il 28 agosto 2014.
- Michael Shermer, Our Neandertal Brethren: Why They Were Not a Separate Species, su scientificamerican.com, 1º luglio 2010. URL consultato il 28 agosto 2014.
- (EN) Neanderthal bone gives DNA clues, su cnn.com.
- (EN) Scientists decode Neanderthal genes, su msnbc.msn.com. URL consultato il 25 maggio 2007 (archiviato dall'url originale il 3 gennaio 2007).
- (EN) Humans and Neanderthals interbred, su cosmosmagazine.com. URL consultato il 25 maggio 2007 (archiviato dall'url originale il 22 febbraio 2009).
- Elizabeth Pennisi, Ancient DNA: No Sex Please, we're Neanderthals, in Science, vol. 316, n. 5827, 18 maggio 2007, p. 967, DOI:10.1126/science.316.5827.967a. URL consultato il 18 maggio 2007.
- Sriram Sankararaman et al., The date of interbreeding between Neanderthals and modern humans, in Plos genetics, vol. 8, n. 10, 1º ottobre 2012, DOI:10.1371/journal.pgen.1002947. URL consultato il 26 novembre 2012.
- Tom Higham, Christopher Bronk Ramsey, Ivor Karavanic, Fred H. Smith e Erik Trinkaus, Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neanderthals, in PNAS, vol. 103, n. 3, 17 gennaio 2006, pp. 553-557.
- (EN) New Machine Sheds Light on DNA of Neanderthals, su nytimes.com, The New York Times.
- (EN) Jason Shogren, R. Horan e E. Bulte, How Trade Saved Humanity from Biological Exclusion: An Economic Theory of Neanderthal Extinction., in Journal of Economic Behavior and Organization, vol. 58, 2005, pp. 1-29.
- Steven L. Kuhn e Mary C. Stiner, What's a Mother to Do? The Division of Labor among Neanderthals and Modern Humans in Eurasia, in Current Anthropology, vol. 47, n. 6, dicembre 2006.
- Cfr. J.E.T. Channell, L. Vigliotti, "The role of geomagnetic field intensity in late Quaternary evolution of humans and large mammals", Reviews of Geophysics, Vol. 57, 29 May 2019 , .
- (EN) Neanderthal exploitation of marine mammals in Gibraltar, in PNAS, n. 105, 22 settembre 2008, pp. 14241-14242, DOI:10.1073/pnas.0807931105.
- (EN) Neanderthal diet like early modern human's, su nhm.ac.uk.
- MP. Richards, PB. Pettitt; E. Trinkaus; FH. Smith; M. Paunović; I. Karavanić, Neanderthal diet at Vindija and Neanderthal predation: the evidence from stable isotopes., in Proc Natl Acad Sci U S A, vol. 97, n. 13, Jun 2000, pp. 7663-6, DOI:10.1073/pnas.120178997, PMID 10852955.
- <Bo Live, magazine dell'Università di Padova, 5 marzo 2020
- Franco Capone, Sequenziato il DNA dell'Homo di neanderthal, su focus.it, Focus, 6.05.2010.
- Da William L. Straus, Jr. e A. J. E. Cave, Quarterly Review of Biology, vol. 32, n. 4, dicembre 1957.
Citato in (EN) Oxford Dictionary of Modern Quotations, Oxford University Press, 2007, ISBN 0-19-920895-6. - A. Leroi-Gourhan 1965, pp. 56-57, figg. 223-225; J. Kozlowski 1992, p. 37 e ss. e figg. 21-25; M. Lorblanchet 1999, p. 203 e ss.
- (FR) Bernard Heuvelmans e Boris F. Porchnev, L'homme de Néanderthal est toujours vivant, in Paris: Plon, 1974.
- G. Giacobini, L. Umley e A. Dem, Les fossiles humains de la Caverna delle Fate (Finale, Ligurie Italienne) et la definition des caracteres Néandertaliens au debut du Würm, in L'Homme de Néandertal, 3: L'Anatomie, Liegi, M. Otte ed E.Trinkaus, 1988, pp. 53-65..
- Museo Archeologico del Finale, su museoarcheofinale.it.
- G. Giacobini e F. D'Errico, La fauna. In Atti Tavola Rotonda Internazionale sull'interpretazione delle impronte umane della Grotta di Toirano, in Rivista di Studi Liguri, vol. 51, n. 4, 1985, pp. 345-352.
- G. Villa, G. Giacobini e Perettoc., thun hohenstein u. 2001, Neanderthal teeth from the Mousterian levels of the Riparo Tagliente (Verona – N-E Italy), in Atti del XIII Congresso degli Antropologi Italiani, Roma, pp. 45-49.. Z
- Grotta di Fumane
- (FR) S. Cotrozzi, Francesco Mallegni e A.M. Radmilli, Fémur d'un enfant néandertalien dans la Buca del Tasso à Metato, Alpi Apuane (Italie), in L'Anthropologie, n. 89, Parigi, 1985, pp. 111-116.
- G. Stefanini, Condizioni fisiche e stratigrafiche della Buca del Tasso, in Archivio per l'Antropologia e l'Etnologia, LII, 1922, pp. 4-9.
- "Neanderthal, dalla Grotta Guattari al Circeo nuove incredibili scoperte" sul sito del Ministero Beni Culturali
- I Paleantropi di Saccopastore di C.A. Blanc (PDF), su quartaer.eu. URL consultato il febbraio 2016.
- Giampiero Dantoni, I livelli musteriani con strumenti su valva di Callista (Callista)chione (L.) nel Salento
- Roberto Filograna, La grotta del cavallo e la sua preistoria
- Arturo Palma di Cesnola, Il Paleolitico della Puglia.
- (EN) Silvana M. Borgognini Tarli, A neanderthal lower molar from Fondo Cattìe (Maglie, Lecce), in Journal of Human Evolution, vol. 12, n. 8, 1983 (archiviato dall'url originale il 3 maggio 2014).
- Silvana Borgognini Tarli, Studio di un molare neanderthaliano (Maglie 1) proveniente dal Fondo Cattie (Maglie, Lecce), in Quaderni; pubblicazione scientifica del Museo di Paleontologia di Maglie, n. 1, 1983.
- Monte Generoso, su montegeneroso.ch. URL consultato il 25 luglio 2011 (archiviato dall'url originale l'8 aprile 2012).
- Parco Curone, Ana, UbiGreen FondazioneCariplo. Relazione archeologica.
- Grosseto, trovate tracce dell'uomo di Neanderthal: «Viveva nella cava di La Pietra»
- Pagina dedicata a "Topolino All'Età Della Pietra" su INDUCKS, su coa.inducks.org, 21 agosto 2014.
Bibliografia
- (EN) Richard E. Green, Anna-Sapfo Malaspinas, Johannes Krause, Adrian W. Briggs, Philip L.F. Johnson, Caroline Uhler, Matthias Meyer, Jeffrey M. Good, Tomislav Maricic, Udo Stenzel, Kay Prüfer, Michael Siebauer, Hernán A. Burbano, Michael Ronan, Jonathan M. Rothberg, Michael Egholm, Pavao Rudan, Dejana Brajković, Željko Kućan, Ivan Gušić, Mårten Wikström, Liisa Laakkonen, Janet Kelso, Montgomery Slatkin e Svante Pääbo, A Complete Neanderthal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing [collegamento interrotto], in Cell, vol. 134, pp. 416-426.
- (EN) Richard E. Green, Johannes Krause, Susan E. Ptak, Adrian W. Briggs, Michael T. Ronan, Jan F. Simons, Lei Du, Michael Egholm, Jonathan M. Rothberg, Maja Paunovic e Svante Pääbo, Analysis of one million base pairs of Neanderthal DNA (PDF), in Nature, n. 444, 16 novembre 2006, pp. 330-336.
- AA. VV., I neanderthaliani. Comparsa e scomparsa di una specie, in Systema Nature, vol. 7, Pavia, Edizioni Altravista, 2009.
- Svante Pääbo L'uomo di Neanderthal, Alla ricerca dei genomi perduti, Einaudi, ISBN 978-88-06-20566-9, 2014.
Voci correlate
- Paleoantropologia
- Preistoria
- Homo heidelbergensis
- Homo sapiens
Altri progetti
 Wikiquote contiene citazioni sull'Homo neanderthalensis
Wikiquote contiene citazioni sull'Homo neanderthalensis Wikimedia Commons contiene immagini o altri file sull'Homo neanderthalensis
Wikimedia Commons contiene immagini o altri file sull'Homo neanderthalensis Wikispecies contiene informazioni sull'Homo neanderthalensis
Wikispecies contiene informazioni sull'Homo neanderthalensis
Collegamenti esterni
- (EN) Homo neanderthalensis, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
- (EN) Homo neanderthalensis, su Fossilworks.org.
- Il Neanderthal non era 'inferiore' al Sapiens, su repubblica.it, la Repubblica, 27 agosto 2008.
- Il Neaderthal si ornava di piume, su repubblica.it, la Repubblica, 25 febbraio 2011.
| Controllo di autorità | Thesaurus BNCF 32365 · LCCN (EN) sh85090493 · GND (DE) 4171356-4 · BNE (ES) XX540103 (data) · BNF (FR) cb12474629x (data) · J9U (EN, HE) 987007563188905171 · NDL (EN, JA) 00568063 |
|---|
На других языках
[es] Homo neanderthalensis
Los neandertales (Homo neanderthalensis, todavía reconocida por algunos investigadores, pero de manera residual, como Homo sapiens neanderthalensis) u hombre de Neandertal es una especie extinta del género Homo que habitó en Europa, Próximo Oriente, Oriente Medio y Asia Central, entre 230 000 y 28 000 años antes del presente, durante el final del Pleistoceno medio y casi todo el superior. Cuando se descubrió, se lo nombró Homo neanderthalensis y se clasificó como una especie distinta del Homo sapiens. No obstante, algunos autores lo consideran una subespecie de Homo sapiens[1] y suelen referirse a dicha subespecie como Homo sapiens neanderthalensis.[2][fr] Homme de Néandertal
- [it] Homo neanderthalensis
[ru] Неандерталец
Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis), в советской литературе также носил название палеоантроп — вымерший или ассимилированный представитель рода людей. Некоторые учёные считают неандертальца подвидом Homo sapiens, другие — отдельным видом рода Homo, поэтому до сих пор сохраняются оба латинских названия. Возраст останков наиболее ранних неандертальцев (пра-неандертальцев) — около 500 тыс. лет[1][2][3][4]; окончательно сформировались, как вид, около 130—150 тыс. лет назад[4]; последние неандертальцы жили около 40 тысяч лет назад[5][6], сосуществовали в Европе с Homo sapiens в течение нескольких тысяч лет. Неандертальцы населяли Европу и Среднюю Азию. Название «неандерталец» происходит от долины Неандерталь в Германии, где был найден череп одного из представителей.Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии