bio.wikisort.org - Animalia
Munychryiinae Common & McFarland,, 1970[1] è una sottofamiglia di lepidotteri, appartenente alla famiglia Anthelidae, endemica dell'Australia.[2][3][4]
| Munychryiinae | |
|---|---|
 | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Sottoregno | Eumetazoa |
| Superphylum | Protostomia |
| Phylum | Arthropoda |
| Subphylum | Tracheata |
| Superclasse | Hexapoda |
| Classe | Insecta |
| Sottoclasse | Pterygota |
| Coorte | Endopterygota |
| Superordine | Oligoneoptera |
| Sezione | Panorpoidea |
| Ordine | Lepidoptera |
| Sottordine | Glossata |
| Infraordine | Heteroneura |
| Divisione | Ditrysia |
| Superfamiglia | Bombycoidea |
| Famiglia | Anthelidae |
| Sottofamiglia | Munychryiinae Common & McFarland, 1970 |
| Generi | |
| |

Descrizione

Si tratta di falene eteroneure appartenenti ai Ditrysia, con apertura alare compresa tra i 22,2 mm di alcuni maschi di Gephyroneura cosmia e i 36,3 mm di alcune femmine di Munychryia periclyta; questi lepidotteri hanno abitudini tendenzialmente notturne o crepuscolari, e sono dotati di una buona capacità di volo. Il corpo tende ad essere alquanto robusto, e in particolare il torace e l'addome appaiono ricoperti da una fitta peluria.[1][2][3][5][6]
Adulto
Capo
Il capo appare rivestito di scaglie piliformi.[5] Non sono presenti ocelli né chaetosemata.[1][2][3][5]
Le antenne hanno in entrambi i sessi una lunghezza leggermente superiore alla metà della costa dell'ala anteriore; nei maschi sono bipettinate fino all'apice, mentre nelle femmine possono essere dentate all'apice, o comunque avere processi laterali che si riducono via via di lunghezza in direzione distale; gli antennomeri hanno un asse centrale rivestito di fitte scaglie.[1][2][3][5]
Le appendici boccali sono meno ridotte di quanto si possa riscontrare nelle Anthelinae. I lobi piliferi sono presenti e muniti di setole laterali più o meno sviluppate. I palpi mascellari sono unisegmentati, mentre quelli labiali sono ricurvi, con l'ultimo segmento piccolo, ascendente e quasi nascosto tra le lunghe setole del secondo. La spirotromba è sempre presente, ma in Munychryia è lunga, arrotolata e munita distalmente di papille carenate, mentre in Gephyroneura è più corta (grosso modo della lunghezza dei palpi labiali), nonché priva di papille distali.[1][2][3][5]
Torace
Il torace è fittamente rivestito di lunghe scaglie piliformi, ma il metascuto appare privo di spinule. Non sono presenti organi timpanici toracici.[1][2][3][5]
Nell'ala anteriore si nota la presenza di un'areola allungata, dal cui apice dipartono le sezioni distali di Rs1, Rs2, Rs3 ed Rs4; nel genere Gephyroneura, la suddetta areola risulta più corta, poiché Rs1, Rs2, Rs3 ed Rs4 si uniscono a livello più prossimale; nella parte distale dell'areola, sono presenti due nervature trasversali: una prima tra Rs1 ed Rs2, oltre a una seconda, posta tra R ed Rs1. M2 corre più ravvicinata a M3 che a M1. CuA1 e CuA2 sono chiaramente separate e distinguibili, mentre manca CuP. 1A+2A mostra una netta biforcazione nella regione basale.[1][2][3][5]
Nell'ala posteriore, provvista di una venulazione decisamente più ridotta, Sc è connessa al margine superiore della cellula discale, in un singolo punto (in Munychryia), oppure tramite una nervatura trasversale di lunghezza molto ridotta (in Gephyroneura). Rs ed M1 sono unite in un solo punto (in Munychryia), oppure per un breve tratto (in Gephyroneura), e come nell'ala anteriore, M2 parte in posizione più ravvicinata a M3 che a M1. Manca CuP, mentre 3A è presente e ben definita.[1][2][3][5]
L'accoppiamento alare è del tipo "frenato" nei maschi e amplessiforme nelle femmine, col mantenimento della sola parte basale del frenulum.[1][2][3] La spinarea è assente[7] e l'apice è appuntito, ma solo lievemente falcato.[1][2][3][5]
Nelle zampe, rivestite di fitte scaglie piliformi, l'epifisi è sempre presente in entrambi i sessi, ma più ridotta nelle femmine, mentre gli speroni tibiali, solitamente non molto lunghi, hanno formula 0-2-2 in Gephyroneura e 0-2-4 in Munychryia; fatta eccezione per quest'ultima, le tibie possono rivelare la presenza di diverse spinule, così come i tarsomeri. Arolio e pulvilli sono sempre presenti, questi ultimi provvisti di un lobo dorsale o di processi setiformi alquanto sviluppati. Le unghie hanno struttura semplice, priva di dentellatura ventrale.[1][2][3][5]
Addome
L'addome è alquanto tozzo in entrambi i sessi e, come il torace, risulta privo di organi timpanici; è rivestito da fitte scaglie piliformi.[1][2][3][5]
Nell'apparato riproduttore maschile, tegumen e vinculum sono uniti nella sclerificazione; le valvae sono alquanto inspessite e arrotondate alla base, con un certo numero di setole distali; lo gnathos è assente, così come i socii; l'uncus è robusto e all'apice assume una forma a spatola o a bulbo, nonché provvisto di fitte setole; l'edeago è affusolato e in alcuni casi provvisto di un cornutus.[1][2][5]
Nel genitale femminile, la bursa copulatrix non è mai molto grande, e sul corpus bursae, più o meno arrotondato o allungato, è presente un signum ricurvo; l'ostium bursae è posizionato nella parte anteriore o centrale della superficie ventrale dell'VIII somite; le apofisi posteriori sono sottili e molto allungate, a differenza di quelle anteriori; le papille anali sono poco sviluppate e provviste di setole.[1][2][5]
Uovo
L'uovo è leggermente appiattito in senso dorso-ventrale, con una forma grosso modo ovoidale; una delle due estremità, lievemente più scura e allargata dell'altra, regge il micropilo. Il chorion appare liscio.[1][2][5]
Le uova vengono deposte singolarmente o a coppie, assicurate al substrato da una sostanza colloidale.[1][5][8]
Larva
La larva di I età presenta poche setole secondarie, essenzialmente sul lobo posteriore delle pseudozampe anali. Al contrario, la larva matura è provvista di numerose setole secondarie, che sono piccole e a forma di spatola. La colorazione generale è verde-grigiastra, con quattro linee longitudinali biancastre per lato, presumibilmente a funzione mimetica. Le pseudozampe appaiono giallastre nei due terzi basali e grigiastre nel terzo distale, coi processi posteriori rosati.[1][2][3][5]
Capo
Il capo, di colore giallo-arancio, ha dimensioni maggiori, se confrontato col resto del corpo, di quanto si possa osservare nelle Anthelinae. L'apparato boccale è di tipo ipognato. Sono presenti sei stemmata per lato, disposti su un arco più o meno regolare. La sutura mediana adfrontale è alquanto lunga.[1][9]
Torace
Nel torace, i segmenti hanno tutti dimensioni alquanto simili. Le zampe protoraciche sono più sviluppate delle altre. Le tibie toraciche rivelano un lobo membranoso distale, che si estende oltre la lunghezza dei tarsomeri. Gli spiracoli del protorace sono più grandi di quelli dell'VIII segmento addominale, che a loro volta superano di poco il diametro di quelli del VII; gli spiracoli degli altri somiti hanno tutti dimensioni più ridotte.[1][8]
Nel protorace, le setole D1, XD1 e XD2 si trovano più o meno su una linea retta verticale, in prossimità del margine anteriore del somite; D2 si trova in posizione posterodorsale rispetto a D1; SD2 è assente; L2 è più breve e in posizione lievemente dorsale rispetto a L1 (che è la setola primaria più lunga in assoluto), ma entrambe sono rette dallo stesso pinaculum; SV1 ed SV2 sono nettamente a forma di spatola, a creare una linea orizzontale; MV2 ed MV3 sono presenti, come pure V1.[1]
Nel meso- e metatorace, D1 è anterodorsale rispetto a D2, ed SD2 è dorsale rispetto a SD1; quest'ultima risulta essere di gran lunga la setola primaria più sviluppata dopo L1 del protorace, e quella con la base più sclerotizzata; L ed SV sono entrambe singole, mentre V1, MD1, MSD1, MSD2 ed MV1 sono tutte presenti.[1]
Addome
Nell'addome si osservano D1 e D2 disposte lungo una linea pressoché orizzontale, ma la seconda si trova in posizione leggermente più dorsale tra i segmenti A1-A7, orizzontale in A8, e più dorsale in A9; SD1 è dorsale e più sviluppata rispetto a SD2 tra A1 e A8, mentre A9 presenta solo una singola setola SD; il gruppo L è a doppia setola tra A1 e A8, con L1 posteroventrale rispetto allo spiracolo ed L2 anteroventrale rispetto a L1; A9 regge solo una setola L; il gruppo SV è a singola setola in A1, a doppia setola in A2 (con SV1 ed SV2 disposte su una linea verticale), assente tra A3 e A6 e a singola setola tra A7 e A9; V1 è presente tra A1 e A9; MD1 si trova solo tra A1 e A8; MV3 è presente tra A1 e A9.[1]
Mentre la larva di I età non ha uncini sulle pseudozampe, quelle dalla II alla V età mostrano via via sempre più uncini; le pseudozampe, che si trovano sui segmenti A3-A6 e A10, sono robuste ma non molto allungate, e munite di 17-19 uncini a completa maturazione; l'ultimo paio di pseudopodi, quello anale, alla V età appare più sviluppato dei precedenti, con 27-29 uncini, oltre che nettamente più appuntito.[1][2] Gli uncini sono robusti e disposti su linee ellittiche incomplete, su un singolo ordine.[1][3][9]
Pupa
La pupa è obtecta, alquanto tozza e ben sclerificata, con le appendici fuse tra loro e col resto del corpo. Capo, torace e appendici sono di una tonalità tra il giallo e l'arancio, mentre l'addome è tinto di un arancio un po' più carico, screziato di rosso-brunastro. Il capo rivela inoltre una coppia di robusti processi anteriori, fortemente sclerificati e ricurvi verso l'interno. I palpi mascellari sono assenti, mentre la spirotromba supera di poco la punta delle ali. Le antenne sono relativamente corte (circa i tre quarti della lunghezza della proboscide). I palpi labiali risultano esposti. L'addome è munito di due soli segmenti mobili, A5 e A6, facilmente distinguibili; ventralmente si notano delle macchie poco definite, che potrebbero rappresentare sorte di cicatrici degli pseudopodi; distalmente termina con una coppia di brevi processi appuntiti e ben sclerificati, ma non sono presenti setole del cremaster.[1][2][5]
Il bozzolo ha forma ellittica e allungata; è opaco, di un bruno pallido, e talvolta incorpora parte delle setole della larva; la pupa non è visibile attraverso la parete.[1][2]
Biologia
Ciclo biologico
Gli adulti sono prevalentemente notturni, con una moderata attrazione per la luce.[2][10]
Nella fase immaginale, la spirotromba è ben sviluppata (soprattutto in Munychryia) e presumibilmente funzionante, a differenza di quanto avviene nelle Anthelinae, sebbene le abitudini alimentari di queste falene siano poco conosciute.[1][5]
Anche le larve sono principalmente attive nelle ore di buio, e non si registra il fenomeno del gregarismo.[3][5]
L'impupamento avviene in un bozzolo fissato alla pianta nutrice.[3][9]

Il bozzolo ha una struttura a doppio strato, essenzialmente costituito da fibre sericee, estruse dalla larva e intessute grazie alla filiera.[3][5]
Alimentazione
Queste larve non sono minatrici fogliari, ma al contrario si alimentano aggrappate alle parti vegetali (principalmente le foglie).[1][2][3][5]
Le uniche piante nutrici riportate in letteratura, appartengono al genere Allocasuarina L.A.S.Johnson, 1982 (Casuarinaceae), come ad esempio:[1][5][8][11]
- Allocasuarina muelleriana (Miq.) L.A.S.Johnson, 1982
- Allocasuarina verticillata (Lam.) L.A.S.Johnson, 1982
Parassitoidismo
Non sono noti casi di parassitoidismo ai danni delle larve di Munychryiinae.[12][13]
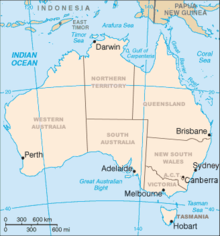
Distribuzione e habitat
Le tre specie appartenenti a questa sottofamiglia sono endemiche dell'Australia: nell'insieme, il taxon è presente in Australia Occidentale, Australia Meridionale, Queensland, Nuovo Galles del Sud, Victoria, Territorio della Capitale Australiana e Tasmania.[1][2][3][5][9]
Tassonomia
Munychryiinae Common & McFarland,, 1970 - J. Austr. Ent. Soc. 9(1): 11[1] - genere tipo: Munychryia Walker, 1865 - List Spec. lepid. Insects Colln Br. Mus. (32): 395.[14]
Generi e specie
La sottofamiglia comprende due soli generi per un totale di tre specie, tutte endemiche dell'Australia:[1][2][5][6][8][10][15][16][17][18]
- Gephyroneura Turner, 1920 - Trans. R. ent. Soc. Lond. 1919(3-4): 417[6] - 1 specie; Australia
- Gephyroneura cosmia Turner, 1921 - Proc. Linn. Soc. N.S.W. 46(2):189[10] - Queensland
- Munychryia Walker, 1865 - List Spec. lepid. Insects Colln Br. Mus. (32): 395[14] - 2 specie; Australia
- = Munychryta [sic] Turner, 1920 - Trans. R. ent. Soc. Lond. 1919(3-4): 417[6]
- = Munichryia [sic] Turner, 1921 - Proc. Linn. Soc. N.S.W. 46(2):189[10]
- Munychryia periclyta Common & McFarland, 1970 - J. Austr. Ent. Soc. 9(1): 19[1] - Australia Occidentale
- Munychryia senicula Walker, 1865 - List Spec. Lepid. Insects Colln Br. Mus. (32): 396[14] - Australia Meridionale, Queensland, Nuovo Galles del Sud, Victoria, Territorio della Capitale Australiana e Tasmania
- = Hypochroma nyssiata Felder & Rogenhofer, 1875 - Reise Fregatte Novara 2 (Abth. 2) (4): tav. 125, fig. 3[19]
Sinonimi
Non sono stati riportati sinonimi.[15][17]
Filogenesi
L'analisi genetica svolta da Heikkila et al. (2015)[20] ha portato alla luce alcune relazioni tra le famiglie di Bombycoidea, così da giungere a un cladogramma da cui è stato ricavato quello riportato qui sotto:
| Bombycoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Anche lo studio pubblicato precedentemente da Zwick et al. (2008) ha confermato la sostanziale monofilia delle due sottofamiglie di Anthelidae.[21]
Conservazione
Nessuna specie appartenente a questa famiglia è stata inserita nella Lista rossa IUCN.[22]
Note
- (EN) Common, I. F. B. & McFarland, N., A new subfamily for Munychryia Walker and Gephyroneura Turner (Lepidoptera: Anthelidae) and the description of a new species from western Australia (PDF), in Journal of the Australian Entomological Society, vol. 9, n. 1, Brisbane, The Society, aprile 1970, pp. 11-22, ISSN 0004-9050, LCCN 76646181, OCLC 4660271339. URL consultato il 10 marzo 2018.
- (EN) Lemaire, C. & Minet, J., The Bombycoidea and their Relatives, in Kristensen, N. P. (Ed.) - Handbuch der Zoologie / Handbook of Zoology, Band 4: Arthropoda - 2. Hälfte: Insecta - Lepidoptera, moths and butterflies, Kükenthal, W. (Ed.), Fischer, M. (Scientific Ed.), Teilband/Part 35: Volume 1: Evolution, systematics, and biogeography, ristampa 2013, Berlino, New York, Walter de Gruyter, 1999 [1998], pp. 321 - 354, ISBN 978-3-11-015704-8, OCLC 174380917. URL consultato il 10 marzo 2018.
- (EN) Scoble, M. J., Higher Ditrysia, in The Lepidoptera: Form, Function and Diversity, seconda edizione, London, Oxford University Press & Natural History Museum, 2011 [1992], pp. 328-341, ISBN 978-0-19-854952-9, LCCN 92004297, OCLC 25282932.
- (EN) Kitching, I. J.; Rougerie, R.; Zwick, A.; Hamilton, C. A.; St. Laurent, R. A.; Naumann, S.; Ballesteros Mejia, L.; Kawahara, A. Y., A global checklist of the Bombycoidea (Insecta: Lepidoptera) (PDF), in Biodiversity Data Journal, vol. 6, Sofia, Pensoft Publishers, 12 febbraio 2018, p. e22236, DOI:10.3897/BDJ.6.e22236, ISSN 1314-2828, LCCN 2014243065, OCLC 967864539. URL consultato il 10 marzo 2018.
- (EN) Common, I. F. B., Moths of Australia, Slater, E. (fotografie), Carlton, Victoria, Melbourne University Press, 1990, pp. vi, 535, 32 con tavv. a colori, ISBN 9780522843262, LCCN 89048654, OCLC 220444217.
- (EN) Turner, A. J., A new family of Lepidoptera, the Anthelidae (PDF), in Transactions of the Royal Entomological Society of London, vol. 67, n. 3-4, Londra, The Society, gennaio 1920, pp. 415-419, DOI:10.1111/j.1365-2311.1920.tb00014.x, ISSN 0035-8894, LCCN sn88024447, OCLC 5156765240. URL consultato il 10 marzo 2018.
- (FR) Minet, J., Nouvelles frontières, géographiques et taxonomiques, pour la famille des CaIlidulidae (Lepidoptera, Calliduloidea), in Nouvelle Revue d'Entomologie (N. S.), vol. 6, n. 4, Fontenay-sous-Bois, Association pour le soutien de la Nouvelle Revue d'Entomologie, 1990 [1989], pp. 351-368, ISSN 0374-9797, OCLC 637361329.
- (EN) Anthelidae, su Butterfly House. URL consultato il 10 marzo 2018.
- (EN) Stehr, F. W. (Ed.), Immature Insects, Vol. 1, seconda edizione, Dubuque, Iowa, Kendall/Hunt Pub. Co., 1991 [1987], pp. 509-524, ISBN 978-0-8403-3702-3, LCCN 85081922, OCLC 13784377.
- (EN) Turner, A. J., Revision of Australian Lepidoptera. Hypsidae, Anthelidae (PDF), in Proceedings of the Linnean Society of New South Wales, vol. 46, Sydney, The Society, 1921, pp. 159-191, DOI:10.5962/bhl.part.14007, ISSN 0370-047X, LCCN 21002639, OCLC 5974844269. URL consultato il 10 marzo 2018.
- (EN) Robinson, G. S.; Ackery, P. R.; Kitching, I. J.; Beccaloni, G. W. & Hernández, L. M., Munychryia, su HOSTS - A Database of the World's Lepidopteran Hostplants, Londra, NHM - Natural History Museum, 2010. URL consultato il 10 marzo 2018.
- (EN) Noyes, J.S., Anthelidae, su Universal Chalcidoidea Database, 2012. URL consultato il 10 marzo 2018.
- (EN) Yu, D. S., Anthelidae, su Home of Ichneumonoidea, 28 aprile 2012. URL consultato il 10 marzo 2018 (archiviato dall'url originale il 5 gennaio 2018).
- (EN, LA) Walker, F., List of the specimens of lepidopterous insects in the collection of the British Museum (PDF), parte 32, Londra, Order of the Trustees, 1865 [1854-66], p. 395, DOI:10.5962/bhl.title.58221, ISBN non esistente, LCCN agr04000350, OCLC 257830843. URL consultato il 10 marzo 2018.
- (EN) Beccaloni G., Scoble M., Kitching I., Simonsen T., Robinson G., Pitkin B., Hine A. & Lyal C., Munychryiinae, su The Global Lepidoptera Names Index, Londra, Natural History Museum, ISSN 2405-8858, OCLC 223993023. URL consultato il 10 marzo 2018.
- (EN) Markku Savela, Munychryiinae, su Funet. URL consultato il 10 marzo 2018.
- (EN) Munychryiinae, su BOLD Systems - Barcode of Life Data Systems, Ontario, Canada, Biodiversity Institute of Ontario, OCLC 73870591. URL consultato il 10 marzo 2018.
- (EN) Munychryiinae, su AFD Australian Faunal Directory, Canberra, Australian Government - Department of the Environment and Energy, 9 ottobre 2008, OCLC 58049221. URL consultato il 10 marzo 2018.
- (DE) Felder, C., Reise der österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859, unter den Befehlen des Commodore Bernhard von Wüllerstorf-Urbair (PDF), parte 9, vol. 2(2), Vienna, Aus der Kaiserlich-Königlichen Hof- und Staatsdruckerei, in Commission bei Karl Gerold's Sohn, 1861, p. tav. 125, fig. 3, DOI:10.5962/bhl.title.1597, ISBN non esistente, OCLC 174025314. URL consultato il 10 marzo 2018.
- (EN) Heikkilä, M., Mutanen, M., Wahlberg, N., Sihvonen, P., Kaila, L., Elusive ditrysian phylogeny: an account of combining systematized morphology with molecular data (Lepidoptera) (PDF), in BMC Evolutionary Biology, vol. 15, n. 1, Londra, BioMed Central, 21 novembre 2015, p. 260, DOI:10.1186/s12862-015-0520-0, ISSN 1471-2148, LCCN 2002243069, OCLC 944275977, PMID 26589618. URL consultato il 17 gennaio 2018 (archiviato dall'url originale il 18 agosto 2017).
- (EN) Zwick, A., Molecular phylogeny of Anthelidae and other bombycoid taxa (Lepidoptera: Bombycoidea) (abstract), in Systematic Entomology, vol. 33, n. 1, Oxford, Blackwell Science, 14 gennaio 2008, pp. 190-209, DOI:10.1111/j.1365-3113.2007.00410.x, ISSN 0307-6970, LCCN 76646885, OCLC 4646424351. URL consultato il 10 marzo 2018.
- (EN) International Union for Conservation of Nature and Natural Resources, IUCN Red List of Threatened Species. Version 2016-3, su IUCN 2016, Cambridge, IUCN Global Species Programme Red List Unit, ISSN 2307-8235, OCLC 943528404. URL consultato il 10 marzo 2018.
Bibliografia
Pubblicazioni
- (EN) Brock, J. P., A contribution towards an understanding of the morphology and phylogeny of the Ditrysian Lepidoptera (abstract), in Journal of natural history, vol. 5, n. 1, Londra, Taylor & Francis, 1º febbraio 1971, pp. 29-102, DOI:10.1080/00222937100770031, ISSN 0022-2933, LCCN 68007383, OCLC 6894673384.
- (EN) Brock, J. P., Pupal protrusion in some bombycoid moths, in Entomologist's Gazette, vol. 41, Faringdon, E.W. Classey, 1990, pp. 91-98, ISSN 0013-8894, LCCN sf93093861, OCLC 704023603.
- (EN) Common, I. F. B. & McFarland, N., A new subfamily for Munychryia Walker and Gephyroneura Turner (Lepidoptera: Anthelidae) and the description of a new species from western Australia (PDF), in Journal of the Australian Entomological Society, vol. 9, n. 1, Brisbane, The Society, aprile 1970, pp. 11-22, ISSN 0004-9050, LCCN 76646181, OCLC 4660271339.
- (EN) Heikkilä, M., Mutanen, M., Wahlberg, N., Sihvonen, P., Kaila, L., Elusive ditrysian phylogeny: an account of combining systematized morphology with molecular data (Lepidoptera) (PDF), in BMC Evolutionary Biology, vol. 15, n. 1, Londra, BioMed Central, 21 novembre 2015, p. 260, DOI:10.1186/s12862-015-0520-0, ISSN 1471-2148, LCCN 2002243069, OCLC 944275977, PMID 26589618. URL consultato il 3 maggio 2019 (archiviato dall'url originale il 18 agosto 2017).
- (EN) Kitching, I. J.; Rougerie, R.; Zwick, A.; Hamilton, C. A.; St. Laurent, R. A.; Naumann, S.; Ballesteros Mejia, L.; Kawahara, A. Y., A global checklist of the Bombycoidea (Insecta: Lepidoptera) (PDF), in Biodiversity Data Journal, vol. 6, Sofia, Pensoft Publishers, 12 febbraio 2018, p. e22236, DOI:10.3897/BDJ.6.e22236, ISSN 1314-2828, LCCN 2014243065, OCLC 967864539.
- (RU) Kuznetzov, V. I. & Stekolnikov, A. A., Comparative and functional morphology of the male genitalia of the Bombycoid moths (Lepidoptera, Papilionomorpha: Lasiocampoidea, Sphingoidea, Bombycoidea) and their systematic position, in Trudy Zoologičeskogo instituta, vol. 134, Leningrado, Akademija nauk SSSR, 1985, pp. 3-48, ISSN 0206-0477, OCLC 715330709.
- (FR) Minet, J., Ébauche d'une classification moderne de l'ordre des Lépidoptères, in Alexanor, vol. 14, n. 7, Parigi, P. André, 1986, pp. 291-313, ISSN 0002-5208, OCLC 3739431.
- (FR) Minet, J., Nouvelles frontières, géographiques et taxonomiques, pour la famille des Callidulidae (Lepidoptera, Calliduloidea), in Nouvelle Revue d'Entomologie (N. S.), vol. 6, n. 4, Fontenay-sous-Bois, Association pour le soutien de la Nouvelle Revue d'Entomologie, 1990 [1989], pp. 351-368, ISSN 0374-9797, OCLC 637361329.
- (EN) Minet, J., Tentative reconstruction of the ditrysian phylogeny (Lepidoptera: Glossata) (abstract), in Entomologica scandinavica, vol. 22, n. 1, Stenstrup, Danimarca, Apollo Books, 1991, pp. 69-95, DOI:10.1163/187631291X00327, ISSN 1399-560X, LCCN 70020995, OCLC 5672447005.
- (EN) Minet, J., The Bombycoidea: phylogeny and higher classification (Lepidoptera: Glossata) (abstract), in Insect Systematics & Evolution, vol. 25, n. 1, Stenstrup, Apollo Books, gennaio 1994, pp. 63-88, DOI:10.1163/187631294x00045, ISSN 1399-560X, LCCN 00252879, OCLC 4637513943.
- (EN) Nakamura, M., Some problems on the classification of moth based on the pupal characters, in Garui tsushin = The Japan heterocerists journal, vol. 60, Tokyo, Japan Heterocerists' Society, 1970, pp. 710-714, ISSN 0286-3537, OCLC 72897796.
- (FR) Niculescu, E. V., Sur les caractères morphologiques utilisés dans la taxonomie et la classification des lépidoptères, in SHILAP Revista de Lepidopterología, vol. 17, n. 65, 1989, pp. 95-108, ISSN 0300-5267, OCLC 162248980.
- (EN) Regier, J. C.; Cook, C. P.; Mitter, C. and Hussey, A., A phylogenetic study of the ‘bombycoid complex’ (Lepidoptera) using five protein-coding nuclear genes, with comments on the problem of macrolepidoteran phylogeny (abstract), in Systematic Entomology, vol. 33, n. 1, Oxford, Blackwell Scientific Publications, gennaio 2008, pp. 175-189, DOI:10.1111/j.1365-3113.2007.00409.x, ISSN 0307-6970, LCCN 76646885, OCLC 225295632.
- (EN) Scott, J. A., On the monophyly of the Macrolepidoptera, including a reassessment of their relationship to Cossoidea and Castnioidea, and a reassignment of Mimallonidae to Pyraloidea (PDF), in Journal of research on the lepidoptera, vol. 25, n. 1, Beverly Hills, CA, Lepidoptera Research Foundation, 1986, pp. 30-38, ISSN 2156-5457, LCCN 2010202002, OCLC 848203568 (archiviato dall'url originale il 15 ottobre 2017).
- (EN) Swinhoe, C., A revision of the Old World Lymantriidae in the National Collection (PDF), in Transactions of the Entomological Society of London, vol. 51, Londra, The Society, 1903, pp. 375-498, ISSN 0035-8894, LCCN sn88024445, OCLC 5156753765.
- (EN) Swinhoe, C., A revision of the genera of the family Liparidae (PDF), in Annals and magazine of natural history; zoology, botany, and geology, (9) 10, n. 58, Londra, Taylor & Francis, 1922, pp. 449-484, ISSN 0374-5481, LCCN 68007383, OCLC 875836246.
- (EN) Turner, A. J., A new family of Lepidoptera, the Anthelidae (PDF), in Transactions of the Royal Entomological Society of London, vol. 67, n. 3-4, Londra, The Society, gennaio 1920, pp. 415-419, DOI:10.1111/j.1365-2311.1920.tb00014.x, ISSN 0035-8894, LCCN sn88024447, OCLC 5156765240.
- (EN) Turner, A. J., Revision of Australian Lepidoptera. Hypsidae, Anthelidae (PDF), in Proceedings of the Linnean Society of New South Wales, vol. 46, Sydney, The Society, 1921, pp. 159-191, DOI:10.5962/bhl.part.14007, ISSN 0370-047X, LCCN 21002639, OCLC 5974844269.
- (EN) Turner, A. J., A review of the phylogeny and classification of the Lepidoptera (PDF), in Proceedings of the Linnean Society of New South Wales, vol. 71, Sydney, The Society, 1947, pp. 303-338, ISSN 0370-047X, LCCN 21002639, OCLC 476489959.
- (EN) Zwick, A., Molecular phylogeny of Anthelidae and other bombycoid taxa (Lepidoptera: Bombycoidea) (abstract), in Systematic Entomology, vol. 33, n. 1, Oxford, Blackwell Science, 14 gennaio 2008, pp. 190-209, DOI:10.1111/j.1365-3113.2007.00410.x, ISSN 0307-6970, LCCN 76646885, OCLC 4646424351.
- (EN) Zwick, A., The principal structure of male genital sclerites and muscles of bombycoid moths, with special reference to Anthelidae (Lepidoptera: Bombycoidea) (abstract), in Arthropod Structure and Development, vol. 38, n. 2, Oxford, Inghilterra, Elsevier Science Ltd., marzo 2009, pp. 147-161, DOI:10.1016/j.asd.2008.07.006, ISSN 1467-8039, LCCN 00260055, OCLC 4933452593, PMID 18765298.
- (EN) Zwick, A.; Regier, J.C.; Mitter, C. and Cummings, M., Increased gene sampling yields robust support for higher-level clades within Bombycoidea (Lepidoptera) (abstract), in Systematic Entomology, vol. 36, n. 1, Oxford, Blackwell Scientific Publications, 1º gennaio 2011, pp. 31-43, DOI:10.1111/j.1365-3113.2010.00543.x, ISSN 0307-6970, LCCN 76646885, OCLC 698219475.
Testi
- (DE) Bryk, F., Lymantriidae, in Lepidopterorum Catalogus, Strand, E., vol. 62, Berlino, Junk, 1934, p. 441, ISBN non esistente, LCCN 12012878, OCLC 630860100.
- (EN) Capinera, J. L. (Ed.), Encyclopedia of Entomology, 4 voll., 2nd Ed., Dordrecht, Springer Science+Business Media B.V., 2008, pp. lxiii + 4346, ISBN 978-1-4020-6242-1, LCCN 2008930112, OCLC 837039413.
- (EN) Common, I. F. B., Australian Moths, Slater, E. (Photos), Brisbane, Jacaranda Press, 1963, pp. 128; 331 figg., ISBN non esistente, LCCN 73028592, OCLC 7401642.
- (EN) Common, I. F. B., Lepidoptera (Moths and butterflies), in CSIRO (ed.). The insects of Australia: a textbook for students and research workers, sponsored by the Division of Entomology, Commonwealth Scientific and Industrial Research Organization, Canberra, I edition, Melbourne, Melbourne University Press, 1970, pp. xiii, 1029, ISBN 9780522838374, LCCN 70461097, OCLC 907783264.
- (EN) Common, I. F. B., Moths of Australia, Slater, E. (fotografie), Carlton, Victoria, Melbourne University Press, 1990, pp. vi, 535, 32 con tavv. a colori, ISBN 9780522843262, LCCN 89048654, OCLC 220444217.
- (DE) Felder, C., Reise der österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859, unter den Befehlen des Commodore Bernhard von Wüllerstorf-Urbair (PDF), parte 9, vol. 2(2), Vienna, Aus der Kaiserlich-Königlichen Hof- und Staatsdruckerei, in Commission bei Karl Gerold's Sohn, 1861, pp. 1-39; tavv. 1-140, DOI:10.5962/bhl.title.1597, ISBN non esistente, OCLC 174025314.
- (EN) Fletcher, D. S. and Nye, I. W. B., The Generic names of moths of the world (PDF), Vol. 4, Bombycoidea, Castnioidea, Cossoidea, Mimallonoidea, Sesioidea, Sphingoidea, Zygaenoidea, Londra, Trustees of the British Museum (Natural History), 1982, p. 192, DOI:10.5962/bhl.title.119597, ISBN 9780565008482, OCLC 55566822.
- (EN) Kristensen, N. P. (Ed.), Handbuch der Zoologie / Handbook of Zoology, Band 4: Arthropoda - 2. Hälfte: Insecta - Lepidoptera, moths and butterflies, Kükenthal, W. (Ed.), Fischer, M. (Scientific Ed.), Teilband/Part 35: Volume 1: Evolution, systematics, and biogeography, ristampa 2013, Berlino, New York, Walter de Gruyter, 1999 [1998], pp. x, 491, ISBN 978-3-11-015704-8, OCLC 174380917.
- (EN) Scoble, M. J., The Lepidoptera: Form, Function and Diversity, seconda edizione, London, Oxford University Press & Natural History Museum, 2011 [1992], pp. xi, 404, ISBN 978-0-19-854952-9, LCCN 92004297, OCLC 25282932.
- (EN) Stehr, F. W. (Ed.), Immature Insects, 2 volumi, seconda edizione, Dubuque, Iowa, Kendall/Hunt Pub. Co., 1991 [1987], pp. ix, 754, ISBN 978-0-8403-3702-3, LCCN 85081922, OCLC 13784377.
- (EN) Swinhoe, C., Catalogue of eastern and Australian Lepidoptera Heterocera in the collection of the Oxford University Museum (PDF), Part I. Sphinges and Bombyces, Oxford, Clarendon Press, 1892, pp. 1-324, ISBN non esistente, OCLC 903998282.
- (EN, LA) Walker, F., List of the specimens of lepidopterous insects in the collection of the British Museum (PDF), parti 31-32, Londra, Order of the Trustees, 1865 [1854-66], pp. 2-706, DOI:10.5962/bhl.title.58221, ISBN non esistente, LCCN agr04000350, OCLC 257830843.
- (EN) Zborowski, P. & Edwards, T., A Guide to Australian Moths, Collingwood, Victoria, CSIRO, 2007, pp. x; 214, ISBN 9780643091597, LCCN 2007408402, OCLC 266969305.
Voci correlate
Altri progetti
 Wikimedia Commons contiene immagini o altri file su Munychryiinae
Wikimedia Commons contiene immagini o altri file su Munychryiinae Wikispecies contiene informazioni su Munychryiinae
Wikispecies contiene informazioni su Munychryiinae
Collegamenti esterni
- (EN) ADW - Animal Diversity Web, su animaldiversity.org. URL consultato il 10 marzo 2018.
- (EN) Aus-lep, su aus-lep.com. URL consultato il 10 marzo 2018.
- (EN) Australian Faunal Directory, su biodiversity.org.au. URL consultato il 10 marzo 2018.
- (EN) BHL - Biodiversity Heritage Library, su biodiversitylibrary.org. URL consultato il 10 marzo 2018.
- (EN) BioLib.cz/, su biolib.cz. URL consultato il 10 marzo 2018.
- (EN) BOLD Systems - Barcode of Life Data Systems, su boldsystems.org. URL consultato il 10 marzo 2018.
- (EN) Butterflyhouse, su lepidoptera.butterflyhouse.com.au. URL consultato il 10 marzo 2018.
- (EN) Catalogue of Life 2017, su catalogueoflife.org. URL consultato il 10 marzo 2018.
- (EN) CSIRO Ecosystem Sciences - Australian Moths Online, su www1.ala.org.au. URL consultato il 10 marzo 2018 (archiviato dall'url originale l'11 marzo 2018).
- (EN) EOL - Encyclopedia of Life, su eol.org. URL consultato il 10 marzo 2018.
- (EN) Flickr, su flickr.com. URL consultato il 10 marzo 2018.
- (EN) Funet.fi, su nic.funet.fi. URL consultato il 10 marzo 2018.
- (EN) GBIF - Global Biodiversity Information Facility, su gbif.org. URL consultato il 10 marzo 2018.
- (EN) Home of Ichneumonoidea, su taxapad.com. URL consultato il 10 marzo 2018 (archiviato dall'url originale l'11 marzo 2018).
- (EN) Insecta.pro/, su insecta.pro. URL consultato il 10 marzo 2018.
- (EN) Internet Archive, su archive.org. URL consultato il 10 marzo 2018.
- (EN) ION - Index to Organism Names, su organismnames.com. URL consultato il 10 marzo 2018.
- (EN) Lepidoptera Barcode of Life, su lepbarcoding.org. URL consultato il 10 marzo 2018.
- (EN) NCBI - National Center for Biotechnology Information, su ncbi.nlm.nih.gov. URL consultato il 10 marzo 2018.
- (EN) NHM Natural History Museum - Butterflies and Moths of the World, su nhm.ac.uk. URL consultato il 10 marzo 2018.
- (EN) NHM Natural History Museum - HOST - a Database of the World's Lepidopteran Hostplants, su nhm.ac.uk. URL consultato il 10 marzo 2018.
- (EN) NHM Natural History Museum - The Global Lepidoptera Names Index, su nhm.ac.uk. URL consultato il 10 marzo 2018.
- (EN) NHM Natural History Museum - Universal Chalcidoidea Database, su nhm.ac.uk. URL consultato il 10 marzo 2018.
- (EN) Nomenclator Zoologicus, su ubio.org. URL consultato il 10 marzo 2018.
- (EN) Pensoft Taxon Profile, su ptp.pensoft.eu. URL consultato il 10 marzo 2018.
- (EN) Systema Naturae - The Taxonomicon, su taxonomicon.taxonomy.nl. URL consultato il 10 marzo 2018.
- (EN) Tree of Life Web Project, su tolweb.org. URL consultato il 10 marzo 2018.
На других языках
[es] Munychryiinae
Los municriínos (Munychryiinae) son una subfamilia de lepidópteros glosados del clado Ditrysia.- [it] Munychryiinae
Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии