bio.wikisort.org - Animalia
Il platecarpo (gen. Platecarpus Cope, 1869) è un rettile marino, appartenente alla famiglia dei mosasauridi. Visse nel Cretaceo superiore (Santoniano - Maastrichtiano, circa 85 - 70 milioni di anni fa), e i suoi resti fossili sono stati rinvenuti principalmente in Nordamerica, ma anche in Europa, Africa e (forse) in Asia e Sudamerica. È noto per numerosi esemplari, tra i quali uno in cui si sono conservati i tessuti molli.
| Platecarpus | |
|---|---|
 | |
| Stato di conservazione | |
Fossile Periodo di fossilizzazione: Cretacico | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Sottoregno | Eumetazoa |
| Superphylum | Deuterostomia |
| Phylum | Chordata |
| Subphylum | Vertebrata |
| Infraphylum | Gnathostomata |
| Superclasse | Tetrapoda |
| Classe | Reptilia |
| Sottoclasse | Diapsida |
| Infraclasse | Lepidosauromorpha |
| Superordine | Lepidosauria |
| Ordine | Squamata |
| Sottordine | Incertae sedis |
| Superfamiglia | † Mosasauroidea |
| Famiglia | † Mosasauridae |
| Sottofamiglia | † Plioplatecarpinae |
| Genere | † Platecarpus Cope, 1869 |
| Specie | |
| |

Descrizione
Questo animale è noto per numerosi esemplari fossili ritrovati principalmente in Nordamerica, nella zona che nel Cretaceo superiore era occupata dal Mare interno occidentale. Come tutti i mosasauri, Platecarpus era dotato di un corpo idrodinamico con quattro zampe trasformate in strutture simili a pinne e una testa triangolare fornita di denti appuntiti e conici. Per lungo tempo si è ritenuto che Platecarpus, come tutti i mosasauri, possedesse una coda appiattita lateralmente e diritta; nuovi studi (Lindgren et al., 2010) hanno ipotizzato la presenza di una coda bilobata, molto simile a quella degli ittiosauri e degli squali. Generalmente Platecarpus era lungo circa 4-5 metri, ma alcuni esemplari potevano raggiungere una lunghezza di oltre 7 metri. Metà della lunghezza totale dell'animale spettava alla coda.
Testa
Il cranio di Platecarpus era insolito se confrontato con quello di altri mosasauri: era corto, con ossa molto mobili, e possedeva meno denti di qualunque altro mosasauro (circa dieci denti su ogni osso dentale). I denti erano lunghi, relativamente sottili e a sezione circolare.

In un esemplare (LACM 128319) si sono conservati numerosi tessuti molli, tra cui alcune tracce di materia all'interno dell'anello sclerotico nell'orbita, che potrebbe essere la retina dell'occhio. Piccole strutture nella retina, ciascuna delle quali lunga 2 µm e osservate grazie alla microspettroscopia, potrebbero rappresentare i melanosomi della retina nelle loro posizioni originali. Attualmente questo esemplare è conservato nel Natural History Museum of Los Angeles County.
Vie respiratorie
Lo stesso esemplare mostra anche il canale respiratorio, conservatosi come anelli cartilaginei (anche se si è conservata solo la parte posteriore della trachea, presso il cinto pettorale). Era conservata anche la sezione dove i due bronchi si dividevano, ma è stata distrutta accidentalmente durante lo scavo. Queste caratteristiche sono un'indicazione del fatto che Platecarpus (e con ogni probabilità anche gli altri mosasauri) possedevano due polmoni funzionali; i serpenti, strettamente imparentati con i mosasauri, ne hanno invece solo uno. Al contrario delle lucertole terrestri, però, i bronchi si separavano davanti all'area delle zampe, e non al livello di queste ultime.
Impronte della pelle

Si sono conservate anche impronte di pelle; le scaglie sulla punta del muso e sulla parte superiore della testa erano esagonali e non venivano a contatto le une con le altre. Quelle delle fauci erano più lunghe e di forma romboidale, e si sovrapponevano fra loro. Le scaglie sul muso indicano che le narici esterne erano posizionate in avanti nel muso, e in posizione laterale come nella maggior parte degli squamati e degli arcosauri. Le scaglie del corpo erano di forma romboidale e formavano file diagonali che si sovrapponevano l'un l'altra al loro margine posteriore. Erano generalmente della stessa misura, ma quelle della coda erano più alte e più grandi (tranne quelle caudali inferiori).
Organi interni

Nell'esemplare LACM 128319 potrebbero essere presenti anche organi interni, conservatisi come aree rossastre. Una di queste è localizzata nella parte bassa della gabbia toracica, mentre l'altra si trova nella porzione superiore della cavità addominale. Le aree rossastre sono state analizzate con uno spettrometro di massa e sono stati trovati alti livelli di composti di pirite e porfirina. Queste sostanze sono la prova di prodotti di decomposizione di emoglobina che potrebbero essersi formati negli organi quando questi si decomposero. Sulla base delle posizioni, l'organo nella gabbia toracica era probabilmente il cuore o il fegato (o entrambi), mentre quello nel ventre era forse un rene, anche se in una posizione più anteriore rispetto ai reni dei varani (i più stretti parenti viventi dei mosasauri). La posizione anteriore dei reni potrebbe essere stata un adattamento verso un corpo più idrodinamico, come avviene nei cetacei.
Sembrerebbe essersi conservato anche parte del tratto digestivo dell'animale, e la zona è riempita di resti di pesci. La forma di questi resti potrebbe delineare la vera forma della parte corrispondente del tratto digestivo, probabilmente il colon. La presenza di scaglie e di ossa non digerite nel colon suggerisce che Platecarpus e altri mosasauri inghiottivano velocemente il cibo, e non lo digerivano e assorbivano completamente nel tratto gastrointestinale. Coproliti del mosasauro Globidens suggeriscono anch'essi una digestione lenta, dal momento che contengono masse di conchiglie frantumate.
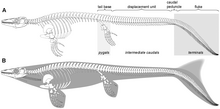
Coda
Le vertebre della coda sono notevolmente incurvate verso il basso. Le vertebre nella zona ricurva (il peduncolo caudale) sono a forma di cuneo, con spine neurali più ampie alle loro estremità che alle loro basi. Questa area ricurva verso il basso probabilmente reggeva un lobo simile a quello degli odierni squali. Potrebbero essere stati presenti due lobi: uno in basso, retto dalle vertebre piegate, e l'altro superiore completamente carnoso. La coda bilobata era probabilmente ipocerca, ovvero con il lobo inferiore più lungo di quello superiore. La stessa condizione si riscontra in altri rettili marini, come gli ittiosauri e i coccodrilli metriorinchidi.

Paleobiologia
Platecarpus era il mosasauro più diffuso nel Mare interno occidentale. In confronto ai mosasauri più grossi (come i tilosaurini), Platecarpus e i suoi stretti parenti possedevano denti meno robusti, ed è probabile che predassero animali più piccoli o teneri, come piccoli pesci o calamari.
Locomozione

Illustrazioni datate di Platecarpus mostrano questo animale con un movimento ondulatorio del corpo mentre nuota, simile a quello delle anguille. Nonostante i mosasauri siano stati tradizionalmente ritenuti rettili anguilliformi, che si spostavano nell'acqua con un'ondulazione laterale del lungo corpo, la profonda pinna caudale di Platecarpus implica che questo animale doveva nuotare in modo simile a quello degli squali. Le vertebre piegate all'ingiù di Platecarpus suggeriscono che possedesse una coda bilobata, e che questa permettesse grande flessibilità in un'area sottoposta a notevoli tensioni durante il nuoto. Le vertebre possiedono inoltre solchi per l'inserzione dei legamenti interspinali e dei tessuti connettivi dorsali, che avrebbero aiutato il movimento laterale dei lobi caudali. I legamenti erano probabilmente costituiti da fibre di collagene che permettevano alla coda di ritornare a una posizione di riposo. Questo tipo di legamenti, in alcuni pesci attuali, serve a conservare energie durante i ripetuti colpi laterali della coda. Mentre i lobi e il dorso della coda si muovevano, la base di essa rimaneva ferma; questo tipo di movimento è noto come locomozione carangiforme (dal nome dei pesci che la utilizzano).

La struttura delle scaglie di Platecarpus potrebbe essere stato un altro adattamento alla vita marina. La piccola taglia e la forma omogenea di queste scaglie potrebbe aver concorso ad irrigidire il corpo, rendendolo più resistente al movimento laterale. Questa rigidità avrebbe migliorato l'efficienza idrodinamica grazie allo scorrimento dell'acqua lungo il corpo. Impronte di scaglie si sono conservate anche sul fossile di un mosasauroide primitivo, Vallecillosaurus, ma queste ultime erano più grandi e di forma variabile, e suggeriscono che l'animale si affidava al movimento ondulatorio del suo corpo piuttosto che esclusivamente su quello della coda. Plotosaurus, un mosasauro più specializzato di Platecarpus, aveva scaglie ancora più piccole, ad indicare un tipo di nuoto ancor più efficiente.
Sindrome da decompressione
Sembra che Platecarpus fosse adattato a una vita marina in acque basse e costiere, e che fosse un predatore d'agguato. Alcune ipotesi, però, vedono Platecarpus come un cacciatore di profondità: già Williston, nel 1898, studiò l'anatomia dell'animale (in particolare l'orecchio esterno ossificato) e pervenne a questa conclusione. Altri studi (Martin e Rotschild, 1989) mostrarono evidenze di sindrome da decompressione (necrosi avascolare) in un'alta percentuale di ossa di Platecarpus; ciò sembrerebbe in accordo con l'idea originaria di Williston. Tuttavia altri studi (Sheldon, 1997) riguardanti l'ispessimento delle costole di questo mosasauro indicherebbero che Platecarpus avrebbe adottato un sistema di caccia simile a quello degli odierni beluga, cacciatori d'agguato in acque basse. Se costretto a scendere in profondità, Platecarpus avrebbe sofferto della sindrome da decompressione.
Tassonomia
I primi resti di questo animale furono scoperti nel Mississippi e vennero descritti da Edward Drinker Cope nel 1869: il paleontologo ascrisse i fossili a un nuovo genere e a una nuova specie di mosasauro, Platecarpus tympaniticus. Il nome generico ("polso piatto") si riferisce alla forma appiattita delle zampe simili a pinne dei mosasauri, in particolare le ossa del polso. L'epiteto specifico, tympaniticus, è in riferimento alle differenze che Cope riscontrò tra l'osso quadrato (allora ritenuto il "timpano") di questo esemplare e quello di Mosasaurus.
Successivamente sono stati scoperti numerosi esemplari di Platecarpus, in particolare nei depositi dello Smoky Hill Chalk in Kansas, attribuite a varie specie: oltre a P. tympaniticus, da ricordare P. ictericus e P. coryphaeus, vissute tra il Santoniano e il Campaniano nel Mare interno occidentale, che nel corso del Cretaceo superiore esisteva nella parte centrale del Nordamerica. Altre specie sono state ritrovate in Angola (P. bocagei, spesso considerata in un genere a sé stante, Angolasaurus) e in Marocco (P. ptychodon, la specie più recente vissuta nel Maastrichtiano). Resti attribuiti a Platecarpus sono stati ritrovati anche in Sudamerica e in Europa. È interessante notare come gli esemplari sudamericani e angolani siano quelli più antichi (Turoniano, circa 88 milioni di anni fa).

La complessa tassonomia del genere Platecarpus ha indotto numerosi studi a tentare una riclassificazione del genere (tra cui Russell 1967, Konishi e Caldell 2007, Konishi et al. 2010); attualmente la specie tipo, P. tympaniticus, comprende anche P. coryphaeus e P. ictericus, tutte risalenti al Santoniano/Campaniano. Un'altra specie attribuita in precedenza a Platecarpus (P. planifrons) e risalente al Coniaciano/Santoniano è attualmente considerata un genere a sé stante (Plesioplatecarpus).
Platecarpus fa parte di una sottofamiglia di mosasauri nota come Plioplatecarpinae, caratterizzata da forme con cranio corto e corpo poco specializzato e relativamente accorciato. Un'altra forma simile è Plioplatecarpus, che differisce dalla precedente principalmente per alcune caratteristiche dei denti e dell'omero.
Bibliografia
- Cope, E. D., 1871, Supplement to the "Synopsis of the Extinct Batrachia and Reptilia of North America.": Proceedings of the American Philosophical Society, v. 12, p. 41-52.
- Williston SW. 1898. Mosasaurs. The University Geological Survey of Kansas, Part V. 4: 81-347 (pls. 10-72).
- Arambourg, C., 1952. Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Service Géologique au Maroc, Notes et Mémoires 92: 1-372.
- Russell, D.A. 1967. Systematics and morphology of American mosasaurs (Reptilia, Sauria). Peabody Museum of Natural History, Yale University, Bulletin, 23:1-241.
- Martin, L. D., and Rothschald, B. M., 1989, Paleopathology and Diving Mosasaurs: American Scientist, v. 77, p. 460-467.
- Lingham-Soliar, T. 1994. The mosasaur "Angolasaurus" bocagei (Reptilia: Mosasauridae) from the Turonian of Angola re-interpreted as the earliest member of the genus Platecarpus. Paläontologische Zeitschrift, 68(1-2):267-282.
- Sheldon, M.A., 1997. Ecological implications of mosasaur bone microstructure. In: Callaway, J.M., & Nicholls, E.L. (eds): Ancient Marine Reptiles. Academic Press (San Diego): 333-354.
- Kiernan, C.R. 2002. Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with an historical review of Alabama mosasaur discoveries. Journal of Vertebrate Paleontology, 22(1):91-103.
- Everhart, M.J. 2003. Revisions to the biostratigraphy of the Mosasauridae (Squamata) in the Smoky Hill Chalk Member of the Niobrara Chalk (Late Cretaceous) of Kansas. Transactions of the Kansas Academy of Science, 104(1-2):59-78.
- Bengtson, P. & Lindgren, J., 2005. First record of the mosasaur Platecarpus Cope, 1869 from South America and its systematic implications. Revista Brasileira de Paleontologia 8: 5-12.
- Konishi, T. and Caldwell, M. W. 2007. New specimens of Platecarpus planifrons (Cope, 1874) (Squamata: Mosasauridae) and a revised taxonomy of the genus: Journal of Vertebrate Paleontology 27(1): 59-72.
- Konishi, T., Caldwell , M.J. and Bell , G.L., Jr. 2010. Redescription of the holotype of Platecarpus tympaniticus Cope 1869 (Mosasauridae: Plioplatecarpinae), and its implications for the alpha taxonomy of the genus. Journal of Vertebrate Paleontology 30(5):1410-1421.
- Lindgren, J.; Caldwell, M.W.; Konishi, T.; and Chiappe, L.M. (2010). "Convergent evolution in aquatic tetrapods: Insights from an exceptional fossil mosasaur". PLoS ONE 5 (8): e11998. doi:10.1371/journal.pone.0011998. PMC 2918493. PMID 20711249. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2918493.
Altri progetti
 Wikimedia Commons contiene immagini o altri file su Platecarpus
Wikimedia Commons contiene immagini o altri file su Platecarpus Wikispecies contiene informazioni su Platecarpus
Wikispecies contiene informazioni su Platecarpus
Collegamenti esterni
Collezione di resti di Platecarpus
На других языках
[es] Platecarpus
Platecarpus ("muñeca plana") es un género extinto de lagarto acuático perteneciente a la familia de los mosasáuridos, que vivió hace cerca de 84-81 millones de años a finales del período Cretácico. Sus fósiles han sido hallados en Estados Unidos.[1] Platecarpus probablemente se alimentaba de peces, calamares y ammonites.[1] Como otros mosasaurios, se pensó inicialmente que nadaba de manera similar a una anguila, aunque un reciente estudio sugiere que nadaba de manera más parecida a los tiburones modernos. Un ejemplar excepcionalmente preservado de P. tympaniticus conocido como LACM 128319 muestra impresiones de piel, pigmentos alrededor de las fosas nasales, tubos bronquiales y la presencia de una alta aleta caudal, mostrando que éste y otros mosasaurios no necesariamente tenían un método de natación similar al de una anguila, más bien eran nadadores rápidos y poderosos. El ejemplar es alojado en el Museo de Historia Natural de Los Ángeles.[2][fr] Platecarpus
Platecarpus est un genre éteint de « reptiles » marins de la famille des mosasaures ayant vécu au Crétacé supérieur. Platecarpus vivait il y a environ 83 millions d'années et a disparu il y a environ 78 millions d'années.- [it] Platecarpus
[ru] Платекарп
Платекарп[источник не указан 1427 дней] (англ. Platecarpus) — вымерший монотипичный род водных ящериц семейства мозазавров, представители которого жили около 85—80 миллионов лет назад в позднемеловом периоде; их ископаемые остатки обнаруживаются в промежутке с «середины» сантонского яруса до «начала» отложений кампанского яруса. Ископаемые остатки были найдены в США, а также возможные образцы (предположительно принадлежащие платекарпу) — в Бельгии и Африке[1]. На данный момент представлен одним видом — Platecarpus tympaniticus.Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии