bio.wikisort.org - Animalia
Los protóstomos o protostomados (Protostomia, del griego «πρῶτος» -prōto- "primero" y «στόμα» -stoma- "boca") son una agrupación de filos del reino animal. Junto con los deuterostomados, forman los dos grandes linajes en que se dividen los animales de simetría bilateral (Bilateria). La separación de estas dos líneas evolutivas fundamentales es ancestral; durante la explosión cámbrica (hace unos 540 millones de años) aparecen ya los filos protostomados y deuterostomados diferenciados.
| Protóstomos | ||
|---|---|---|
|
Rango temporal: Ediacárico – Reciente PreЄ
Є
O
S
D
C
P
T
J
K
Pg
N
| ||
 Protóstomos | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Subreino: | Eumetazoa | |
| (sin rango) | Bilateria | |
| (sin rango): |
Protostomia Grobben, 1908 | |
| Superfilos y filos | ||
| Sinonimia | ||
| ||
La diferencia entre ambos clados, propuesta por Grobben,[1] radica en el origen embrionario de la boca.[2]En los protostomados, la boca del adulto deriva del blastoporo embrionario (de ahí proviene el nombre del grupo), mientras que en los deuterostomados es de neoformación. Sin embargo, en los últimos años se ha visto que el destino del blastoporo y el origen del ano es mucho más complejo y variable entre los grupos de protostomados.
Definición y apomorfías
Karl Grobben, biólogo austríaco, fue quien acuño los términos Protostomia (Urmundtiere) y Deuterostomia (Neumundtiere), los cuales aparecen por primera vez en Die systematische Einteilung des Tierreiches.[1] En este libro, el austríaco agrupó dentro de los protostomados a todos aquellos animales en donde la boca adulta se origina a partir del blastoporo embrionario. Si bien el concepto de clasificación propuesto por Grobben (destino del blastoporo) era nuevo, no lo era el grupo, ya que anteriormente Hatschek[3] había agrupado los mismos taxones bajo el ensamble Zygoneura. La característica que unía a los Zygoneura es que en todos ellos el cordón nervioso era ventral y par. Hoy en día, estos dos caracteres son considerados como apomorfías morfológicas del clado Protostomia.
En la actualidad, ya casi no cabe duda de que los protostomados forman una unidad evolutiva. Las principales apomorfías que sustentan a este grupo son:[4]
- Formación simultánea de la boca y ano a partir del blastoporo, condición conocida como anfistomía (contraposición a Grobben).
- Sistema nervioso formado por un cerebro dorsal —circunfaringeo—, y un par de cordones nerviosos ventrales, a veces fusionados (zigoneuría de Hatschek).
- Larva primaria tipo trocófora, con un sistema ciliar de recolección del tipo downstream (la larva primaria falta en los Ecdysozoa, que carecen de cilias).
- Las primeras dos divisiones del embrión originan una célula (D) que tiene un destino especial, contribuyendo a la formación de gran parte del ectodermo, endomesodermo y células germinales.
Destino del blastoporo
El nombre protostomados deriva de que la boca se forma a partir del blastoporo, por contraposición a los deuterostomados, en los que el blastoporo origina solamente el ano. Sin embargo, el destino del blastoporo no es único y para algunos grupos se dio por sabido aunque no se había estudiado específicamente.
Cuando Martín-Durán et al.[5] estudiaron el desarrollo embrionario de Priapulus caudatus (Priapulida), un protostomado, comprobaron que esta especie presentaba deuterostomía, es decir, el blastoporo da origen al ano, similar a lo que pasa en los deuterostomados. Por otro lado, en algunos grupos (no en todos, pero sí en los más numerosos, como Annelida, Arthropoda y Nematoda), los labios laterales del blastoporo se fusionan, dejando dos aberturas que se convierten en la boca y el ano: presentan anfistomía. En el caso de Polygordius, el orificio posterior se cierra completamente y el ano aparece más tarde, por una invaginación de la pared del cuerpo. Por último, en algunos grupos tales como los picnogónidos el blastoporo origina únicamente la boca, en este caso hablamos de protostomía sensu stricto.
En resumen, a diferencia de la concepción clásica de que los protostomados presentan únicamente protostomía, la realidad es que el tipo de desarrollo varía de acuerdo al grupo en cuestión, pudiendo ser por protostomía, deuterostomía o comúnmente anfistomía (esta última situación es considerada apomórfica para Nielsen[4] y otros autores).
Viendo este panorama, los científicos pasaron a preguntarse cuál sería entonces el carácter plesiomórfico de los Protostomia: ¿protostomía o deuterostomía? Martín-Durán et al.[5] concluyeron que:
- La expresión de los genes bra, cdx, gsc y foxA confirman la homología de la boca y el ano entre los distintos phyla y que la condición ancestral de los Ecdysozoa es la deuterostomía.
- Se sostiene la hipótesis de que el ancestro común entre los Deuterostomia y los Protostomia tenía un desarrollo de tipo deuterostomado (en cuanto al destino de su blastoporo). Esto está en oposición a las teorías clásicas de que la protostomía sería la condición ancestral.
Desarrollo embrionario

Como norma general, los Protostomia tienen desarrollo determinado, a partir de huevos en mosaico. Forman la boca a partir del blastoporo, por protostomía (solo la boca), anfistomía (boca y ano al mismo tiempo) o por deuterostomía (primero se forma el ano a partir del blastoporo y luego la boca en el extremo opuesto del animal, por ejemplo en Priapulida).
Pueden tener segmentación del huevo en T (como en Nematoda y Rotifera), o radial (como en Ectoprocta) o en espiral (Platyhelminthes, Annelida, Nemertea, Mollusca).
En el estadio embrionario de 4 células, una de ellas (célula D) se diferencia del resto y da origen al endomesodermo y a las células germinales. Nielsen[4] nombra a este patrón «segmentación en cuadrante», y lo propone como una sinapomorfía de los Protostomia.
La «célula» D (y su destino) se ha podido identificar en los Spiralia, en el molusco Sphaerium, en el rotífero Asplanchna, en la mayoría de los Ecdysozoa (artrópodos y nematodes) y en los Annelida. Los Brachiopoda y Phoronida tienen patrones de segmentación similares a los Deuterostomia.
Segmentación espiral
En este tipo de segmentación del huevo fecundado, las dos primeras divisiones son longitudinales y perpendiculares entre sí, mientras que las divisiones subsiguientes son oblicuas. Como resultado se forman células hijas de distintos tamaño: macrómeras y micrómeras.
Las nuevas micrómeras que se van formando se desplazan lateralmente en sentido horario (dextrotrópico) o antihorario (levotrópico). El pasaje de 4 a 8 células implica un desplazamiento dextrotrópico. Las siguientes divisiones alternan entre ambos tipos de desplazamientos.[4][6]
Wilson[7] elaboró un elegante sistema de codificación para la segmentación espiral (Sistema de Wilson), el cual permite seguir el linaje de cada célula embrionaria:
- En el embrión de «4 células», éstas son codificadas como A, B, C y D, siguiendo el sentido de las agujas del reloj (vistas desde el polo animal).

- Estas 4 células definen un cuarteto de macrómeras, y delimitan 4 cuadrantes (de ahí el nombre propuesto por Nielsen de segmentación en cuadrantes). Estas células colectivamente son codificadas genéricamente como Q.




- La siguiente división (4 → 8) es desigual y oblicua, generándose macrómeras inferiores y micrómeras superiores, estas últimas se desplazan en forma dextrotrópica. Éstas forman el primer cuarteto de micrómeras (1q): 1a, 1b, 1c y 1d. Ahora, las macrómeras (que siempre permanecen en su lugar, en esta y las divisiones siguientes) se designan como 1Q: 1A, 1B, 1C y 1D. Es decir, cada una de las 4 célula (por ejemplo la A) se divide dando origen a una micrómera (1a) y una macrómera (1A).

- El paso de 8 a 16 células implica la división tanto de macrómeras (1Q) como de micrómeras (1q), y las nuevas micrómeras formadas se desplazan en sentido levotrópico:
- Las macrómeras (1Q) se dividen produciendo un segundo cuarteto de micrómeras (2q), y una segunda serie de macrómeras in situ (2Q).
- Las micrómeras (1q) también se dividen, produciendo nuevas micrómeras que se identifican con un subíndice (1qx).
- El pasaje de 16 a 32 células nuevamente conlleva a un desplazamiento dextrotrópico de las micrómeras hijas:
- Las macrómeras (2Q) vuelven a dividirse para formar un tercer cuarteto de micrómeras (3q), formándose nuevas macrómeras (3Q) in situ.
- Las 12 micrómeras (2qx) también vuelven a dividirse, de esta manera, según el sistema de Wilson, el subíndice queda definido por 2 dígitos (2qxy). De esta manera, de la micrómera 1a1 surgen las células 1a11 y 1a12.
- La división siguiente (32 → 64) ocurre en un patrón levotrópico, dando como resultado hasta el momento las siguientes células:
Generalmente, a partir del embrión de 64 células la segmentación deja de ser espiral y adquiere distintos patrones según los grupos. Como se puede ver, en cada división las nuevas células van rotando de derecha a izquierda, distribuyéndose en un patrón similar a una espiral, de donde deriva el nombre de este tipo de segmentación.

Más tarde en el desarrollo se pueden observar distintos patrones característicos como consecuencia de la orientación del primer cuarteto de micrómeras.
- Las células más superiores yacen en el ápice del embrión (derivados de 1q111) formando la roseta apical.
- En los anélidos y otros grupos las micrómeras derivadas de 1q112 forman la cruz anélida (Mollusca: roseta periférica), perpendicular a la roseta apical.
- En los moluscos aparece una cruz molusca formada a partir de los derivados de la micrómera 1q12.
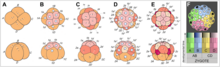
Los Platyhelminthes, Nemertea y el clado Schizocoelia tienen un modelo muy uniforme de segmentación espiral. Hay algunas diferencias en la disposición de las células que forman una "cruz" en el polo animal del embrión (por ejemplo, entre Platyhelminthes y Annelida), pero el modelo general es el mismo.
Origen de las capas germinales


Durante la tercera división del embrión ocurre la separación entre las células que originarán el ectodermo y el endodermo.[6] El cuarteto de micrómeras (q) forman el tejido presuntivo que originará el ectodermo: el «ectoblasto». Las macrómeras (Q) originan el endodermo presuntivo: el «endoblasto».
La macrómera 3D se divide originando la micrómera 4d, la cual se dividirá y formará el mesodermo presuntivo: el «mesoblasto». Esta micrómera (4d) recibe el nombre de mesentoblasto y origina el mesodermo en todos los animales con segmentación espiral. Al dividirse, el mesentoblasto origina dos teloblastos (4d1 y 4d2), que por mitosis originan dos bandas de tejido mesodérmico en la zona de contacto entre el ectodermo y endodermo, cerca del blastoporo.
La gastrulación varía de acuerdo al grupo en cuestión y la cantidad de vitelo que trae el huevo:
- Huevos con poco vitelo (microlecitos) dan origen a una celoblástula, y la gastrulación ocurre mayormente por invaginación.
- Huevos con mucho vitelo (telolecitos, macrolecitos) dan origen a una estereoblástula, y la gastrulación ocurre mayormente por epibolia.
Posterior a la formación del mesodermo ocurre la formación del celoma, cuando este existe. En este caso, y particularmente en el clado Schizocoelia el celoma se forma por ahuecamiento del mesodermo, proceso conocido como esquizocelia: se forma un esquizoceloma.
Larva trocófora
Durante el estadio final de desarrollo embrionario, el embrión puede transformarse en una larva (desarrollo indirecto: esta larva puede ser larva trocófora o no-trocófora) o desarrollarse directamente en un juvenil (desarrollo directo). Ya desde los estudios llevados a cabo por Hatschek[8] se planteó la presencia de larva trocófora como una característica de todos los Protostomia. Sin embargo, no todos los protostomia poseen este tipo de larva. Se pueden plantear tres grandes generalidades acerca de la larva trocófora:
- Es característica de todos los integrantes del clado Schizocoelia o Trochozoa.
- Está presente en otros phyla, considerada como larva trocófora modificada, ya que su morfología difiere del modelo básico.
- Los Ecdysozoa carecen de trocófora, como resultado de una pérdida secundaria.
En la siguiente tabla[4] se muestran los phyla con larva trocófora [+], trocófora modificada [(+)], sin larva trocófora [-] y sin información disponible [?] :
| Phylum | Larva trocófora |
|---|---|
| Annelida | + |
| Mollusca | + |
| Nemertea | + |
| Platyhelminthes | (+) |
| Gastrotricha | ? |
| Gnathostomulida | ? |
| Micrognathozoa | ? |
| Rotifera | (+) |
| Entoprocta | + |
| Cycliophora | (+) |
| Ectoprocta | - |
| Phoronida | - |
| Brachiopoda | - |
| Onychophora | - |
| Arthropoda | - |
| Tardigrada | - |
| Nematoda | - |
| Nematomorpha | - |
| Priapulida | - |
| Kinorhyncha | - |
| Loricifera | - |
| Chaetognatha | - |

Morfológicamente, la larva trocófora se caracteriza por tener forma de peonza o trompo (de ahí deriva su nombre) y por estar surcada por bandas ciliadas. Puede ser lecitotrófica o planctotrófica, dependiendo del desarrollo embrionario, y se mueve gracias a la acción de la bandas ciliadas, que en el caso de las larvas planctotróficas también participan en la alimentación. El cuerpo se divide en dos porciones, separadas por el prototroco (banda ciliar): una semiesfera superior (episfera) y una semiesfera inferior (hiposfera).[4][9][10][11]
- A nivel de la episfera se origina el sistema nervioso de la larva, formado por un órgano apical, un par de ganglios cerebrales y un par de cordones nerviosos ventrales más o menos fusionados (estos últimos se continúan a nivel de la hiposfera). Del órgano apical (placa sincipital) parten un penacho de cilias (penacho apical) característico de esta larva.
- A nivel de la hiposfera se extienden la mayoría de las bandas ciliadas y es aquí donde se evidencias algunos órganos: tubo digestivo (con boca central y ventral, y ano terminal e inferior) y protonefridios (2).
La cavidad interna está representada por el blastocel embrionario. Distintivo de la larva trocófora es la presencia de cinco bandas ciliadas: prototroco (supraoral), metatroco (suboral), zona ciliada adoral (circumoral), gastrotroco (conecta metatroco con telotroco) y telotroco (supraanal).
- El «prototroco» y el «telotroco» (cuando está presente) forman el sistema locomotor de la larva.
- El «prototroco», la «zona ciliada adoral» y el «metatroco» forman el sistema de alimentación de la larva, un sistema de recolección downstream.
- En las trocóforas lecitotróficas falta el «metatroco» y la «zona ciliada adoral» está poco desarrollada.
- En las larvas de algunas especies pueden aparecer bandas ciliadas accesorias («acrotroco», «meniscotroco», «pretroco»).
Valor filogenético de la segmentación espiral
Sobre la base de la segmentación espiral se fundamentó el clado Spiralia, que reunía -como grupos principales- a los Platyhelminthes, Nemertea, Annelida y Mollusca, que siguen el modelo espiral típico.[4]
Si todos los huevos en mosaico, con desarrollo determinado, derivaran de un ancestro con segmentación espiral, se justificaría incluir en un solo clado los grupos con otros tipos de segmentación que tradicionalmente se interpretaron como espiral modificada (aunque a veces no tengan mucha semejanza con el supuesto modelo "original").
Los Spiralia (o algunos de ellos) también recibieron el nombre de Trochozoa, dentro de los cuales distintos autores incluyeron más o menos grupos, según las evidencias que tenían.
Cuando se propuso que los lofoforados (Brachiopoda, Phoronida) eran protostomados, se los unió a los Trochozoa anteriores, con el nombre de Lophotrochozoa. A partir de entonces se discutió la monofilia de los Lophotrochozoa, que a veces se restringe a los Spiralia sensu stricto y a veces incorporan a otros filos. Como no hay un mínimo de acuerdo sobre la filogenia de estos grupos, por ahora es preferible no usar los conceptos Lophotrochozoa y Spiralia, aunque probablemente este último puede tener mayor fundamento.
Celoma
El mesodermo de los protostomados surge como un tejido macizo que crece como dos masas laterales al blastoporo y usualmente prolifera tapizando por dentro al ectodermo. A medida que se desarrolla puede:
- Generar distintos tejidos y órganos en la cavidad blastocélica (animales pseudocelomados).
- Producir un parénquima que rellena el blastocel (animales acelomados o parenquimios).
- Formar dos masas que se ahuecan, originando una cavidad tapizada por peritoneo (animales esquizocelomados).
Cuando se forma una cavidad del cuerpo, el mesodermo que lo rodea puede diferenciar:
- La dermis (un tejido conectivo mesodérmico que se integra al epitelio): puede ser gruesa, delgada o faltar (la mayoría de los invertebrados no tienen dermis).
- Los músculos de la pared del cuerpo (típicamente una capa de múculo circular y, por debajo, una de músculo longitudinal).
- El peritoneo (un epitelio interno, con membrana basal, que a veces se puede perder total o parcialmente durante la ontogenia o bien -en varios phyla de animales con cavidad del cuerpo- no formarse nunca).
| Filo | Acelomado | Pseudocelomado | Celomado |
|---|---|---|---|
| Annelida | |||
| Mollusca | |||
| Nemertea | |||
| Platyhelminthes | |||
| Gastrotricha | |||
| Gnathostomulida | |||
| Micrognathozoa | |||
| Rotifera | |||
| Entoprocta | |||
| Cycliophora | |||
| Ectoprocta | |||
| Phoronida | |||
| Brachiopoda | |||
| Onychophora | |||
| Arthropoda | |||
| Tardigrada | |||
| Nematoda | |||
| Nematomorpha | |||
| Priapulida | |||
| Kinorhyncha | |||
| Loricifera | |||
| Chaetognatha | |||
| Orthonectida |
Tradicionalmente se llama acelomados a los animales parenquimios (sin cavidad del cuerpo), pseudocelomados a los que poseen una cavidad del cuerpo que no esté tapizada por peritoneo y celomados a los que tienen peritoneo. Son conceptos meramente descriptivos, sin ninguna connotación evolutiva.
Animales acelomados
En un acelomado el mesodermo forma un parénquima que ocupa la mayor parte del espacio interno del cuerpo. Sin embargo, no son totalmente macizos, porque dentro del parénquima se abren lagunas en las que se acumula líquido intersticial y quedan con pequeñas cavidades que rodean a las gónadas u otras estructuras. Un claro ejemplo de esto lo son las especies pertenecientes al filo Platyhelminthes.
Animales pseudocelomados
Pseudocelomado es cualquier animal cuya cavidad general del cuerpo no está cubierta por peritoneo (sin importar por qué no lo tiene): no es un concepto con valor filogenético. Antiguamente formaban el filo Aschelminthes (hoy obsoleto), pero dado que el pseudocel no es homólogo en todos los casos, no puede considerarse a los pseudocelomados como un único filo por compartir ese carácter. El pseudoceloma puede surgir por dos mecanismos diferentes, dependiendo del grupo en cuestión:
- Animales con estereogástrula. En estos animales el pseudoceloma se forma por ahuecamiento de un parénquima sincicial (Rotifera, Gnathostomulida).
- Animales con celogástrula. En estos animales el pseudoceloma es el blastocel, que persiste en el adulto (Nematoda, Nematomorpha, Kinorhyncha, Priapulida, Loricifera).

Un caso particular son los gastrotricos, los cuales no tienen cavidad del cuerpo. Sin embargo, muchos científicos los consideran pseudocelomados en donde el pseudoceloma desapareció como consecuencia de una invasión de parénquima mesodérmico (son acelomados funcionales).
En muchas especies como los nematodos, el pseudoceloma no es una cavidad vacía: contiene líquido que funciona como esqueleto hidrostático y siempre se encuentra parcialmente ocupado por grandes células (pseudocelomocitos) de función poco comprendida. Mecánicamente los pseudocelomocitos contribuyen a que los órganos ocupen posiciones relativamente estables.
Animales celomados
En un celomado la cavidad general del cuerpo está envuelta por peritoneo, que se integra a la pared del cuerpo como una somatopleura, rodea al intestino como esplacnopleura, y forma los mesenterios que unen a ambas pleuras. Ejemplo de animales celomados son los gusanos segmentados Annelida.
![Distribución de quitina en los animales según Willmer.[12]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/4c/Quitina_en_animales.png/220px-Quitina_en_animales.png)
Quitina

La quitina es un hidrato de carbono, polímero de N-acetilglucosamina (un azúcar derivado en el que un grupo hidroxilo [-OH] está sustituido por un grupo amino [-NH2]. Sus monómeros se unen entre sí con enlaces β-1,4, igual que las moléculas de glucosa que componen la celulosa. Es el polímero natural más abundante después de la celulosa. También está presente en los hongos.
Se ha detectado la presencia de quitina en al menos 21 filos de animales (Willmer).[12] La mayoría (19) pertenecen al clado Protostomia, y dos de ello son taxones más basales (esponjas y ctenóforos).
Dado que existe quitina en la mayoría de los filos animales y en los otros grupos (hongos), y que está ausente únicamente en los Deuterostomia, la hipótesis más parsimoniosa es pensar que la quitina es una sinapomorfía de los animales. Por lo tanto, se deduce que en los Protostomia la quitina es plesiomórfica, mientras que en los Deuterostomia es apomórfica su ausencia.
Sistema excretor
La excreción es el proceso de eliminación de los desechos nitrogenados que provienen del metabolismo celular. Involucra tres etapas: ultrafiltración, reabsorción y secreción de orina. Se lleva a cabo mediante distintas estructuras, según el phylum: protonefridios, metanefridios, nefridios glandulares, nefridios sacciformes (artrópodos), glomérulos u otros órganos.
El tipo de nefridios está relacionado con el tipo y amplitud de la cavidad general del cuerpo del animal. Así, suelen carecer de sistema excretor (pero no de excreción) los animales más pequeños (menores a 1 mm) y los gelatinosos.
Protonefridios
Un protonefridio es una estructura derivada del ectodermo, que cumple por sí misma los tres procesos de excreción: ultrafiltración, reabsorción y secreción.
Es un tubo cerrado en su extremo por una célula terminal, que tiene uno o más undulipodios (cilias/flagelos) especiales, que mueven el líquido (orina en formación) a lo largo de un tubo que desemboca al exterior por un nefroporo.
Los animales más pequeños y las larvas suelen tener solo dos protonefridios, que en general desembocan juntos en un poso poro. En los animales más grandes, puede haber muchos protonefridios (a veces, miles) que unen sus conductos en dos grandes nefridioductos, que pueden desembocar en una vejiga urinaria.
Cuando sólo hay uno o dos undulipodios, se dice que es un protonefridio con solenocitos; cuando la célula terminal tiene un mechón de cilias, se dice que es un protonefridio con célula flamígera. La diferencia no es absoluta y, en general, solo se dice que un organismo tiene protonefridios, salvo que sea un ejemplo típico de uno de esos dos modelos.
La célula terminal está envuelta por una membrana basal, porosa, a través de la cual se produce la ultrafiltración del líquido de la cavidad (o de intersticios del parénquima si no hay cavidad). El movimiento del o de los undulipodios es lo que produce una presión negativa (succión) que alcanza para que las moléculas pequeñas pasen al interior del protonefridio. A lo largo del nefridioducto se produce la reabsorción de sustancias metabólicamente aprovechables, que son devueltas al celoma (y a su vez se secretan sustancias de desecho al tubo, ayudando a la formación de la orina final).
Metanefridios

Los metanefridios se diferencian de un protonefridio porque:
- Son de origen mesodérmico.
- No realizan la ultrafiltración (esta se hace en podocitos asociados al sistema circulatorio).
- Están abiertos al celoma por una boca ciliada, generalmente en forma de embudo (nefrostoma ciliado).
Los podocitos son células que tienen prolongaciones similares a dedos (pedicelos), asociados a vasos sanguíneos. La presión sanguínea hace que las moléculas pequeñas y positivas pasen el líquido celómico, atravesando la membrana basal.
Los metanefridios toman el líquido celómico y, mediante reabsorción y secreción a lo largo del nefridioducto, producen la orina, que es transportada hasta el nefroporo.
¿Protonefridios o metanefridios?
La existencia de metanefridios supone un sistema circulatorio con podocitos: no hay metanefridios en animales acelomados o pseudocelomados, donde no hay un peritoneo.
Las larvas de los celomados desarrollan protonefridios, de origen ectodérmico, antes de que completen la formación de la cavidad general del cuerpo y del sistema circulatorio. En los celomados, los protonefridios larvales desaparecen durante la metamorfosis y se forman los metanefridios. Solo por excepción los protonefridios larvales permanecen en el adulto.
Los animales que tienen el celoma muy reducido modifican los metanefridios, originando estructuras de funcionamiento diferente: por ejemplo, moluscos (órgano de Bojanus) y artrópodos (tubos de Malpighi).
Tienen protonefridios los adultos de Platyhelminthes, Nemertea, Rotifera, Priapulida, Kinorhyncha, Loricifera, Gnathostomulida, Entoprocta, y las larvas de los esquizocelomados (moluscos y anélidos), Phoronida, Ectoprocta y Gastrotricha.

No tienen sistema excretor los adultos de Nematomorpha, Ectoprocta, Cycliophora.
Nefromixia
Se denomina nefromixia al uso de los conductos del sistema excretor para la emisión de las gametas que maduran libres en el celoma. Puede ocurrir tanto en animales con protonefridios (protonefromixia) como en animales con metanefridios (metanefromixia). La fecundación en ambos casos es generalmente externa (ovulíparos), pero también hay casos de fecundación interna (ovíparos), sin cópula y sin incubaciones largas.
Considerando esto y la relación entre sistemas excretor y reproductor, se puede dar lo siguiente:
- Gonoducto separado del protonefridio; cada uno abre en un poro independiente (gonoporo y nefroporo).
- Fusión del embudo del gonoducto con el conducto del protonefridio; un solo poro permite la emisión de gametas y la salida de la orina.
- Gonoducto separado del metanefridio; dos poros.
- Fusión del embudo del gonoducto delante del embudo metanefridial.
Sistemática y filogenia
Tradicionalmente se llama acelomados a los animales parenquimios (sin cavidad del cuerpo), pseudocelomados a los que poseen una cavidad del cuerpo que no está tapizada por peritoneo y celomados a los que tienen peritoneo. Antiguamente, la sistemática tradicional del siglo XX le dio a estos conceptos un sentido evolutivo, considerándolos grupos naturales:
- Acelomados: Platyhelminthes y Nemertea.
- Pseudocelomados: Nematoda, Rotifera, Priapulida, Gastrotricha, etc.
- Celomados: Annelida, Arthropoda, Mollusca, Bryozoa, etc.
Se asumía una secuencia filogenética: acelomado → pseudocelomado → celomado. Este concepto evolutivo de celomía está totalmente obsoleto, porque hoy se sabe que:
- En algunos celomados, secundariamente, la cavidad del cuerpo se llena de parénquima: son acelomados funcionales (por ejemplo: caracoles, sanguijuelas).
- El pseudoceloma es homoplásico: phyla con distinto origen filogenético carecen de peritoneo; otros son celomados que lo han perdido y en otros la cavidad es una neoformación.
- El peritoneo es homoplásico: apareció al menos dos veces en la evolución, por dos mecanismos ontogenéticos diferentes (en los deuterostomados el peritoneo deriva del endodermo).
En consecuencia, gracias a los análisis cladísticos a partir de datos morfológicos y moleculares, esta visión clásica y errónea de los protostomados empezó a cambiar. Las relaciones entre anélidos y artrópodos (Articulata) han sido invalidadas y ha surgido la hipótesis de los Ecdysozoa, según la cual, los artrópodos estarían emparentados con grupos pseudocelomados que mudan la cutícula. La nueva sistemática filogenética divide a los protostomados en los clados Ecdysozoa y Spiralia:[4]
- Ecdysozoa: incluye a animales que tienen una cutícula de material orgánico (quitina; colágeno) de tres o más capas, que se renueva periódicamente por mudas (ecdisis) para lo que se sintetiza una enzima específica (ecdisona). A su vez carecen de cilias locomotoras, lo que se traduce en la formación de espermatozoides ameboides.
- Spiralia: Incluye a animales que tienen segmentación espiral del huevo, sin embargo no todos sus miembros poseen esta característica, pero se ha sugerido que estos últimos pudieron evolucionar de ancestros que tenían este patrón de desarrollo embrionario. Incluye filos muy diversos morfológicamente que esta respaldado por todos los análisis moleculares. Sus miembros pueden tener como sinapomorfías evolutivas larvas trocóforas.[13]
Árbol filogenético
La filogenia de los protótosmos es muy difícil de resolver, los análisis moleculares dan resultados muy diferentes con la colocación de los grupos. Aunque todo depende del método que se utilice en el análisis. Actualmente filos como Platyhelminthes, Tardigrada, Rotifera tienen tasas de evolución rápida lo que impide reconstruir un árbol filogenético adecuado y llevan a la atracción de ramas largas. Estudios filogenéticos recientes (2019) utilizando especies de evolución más lenta que son menos proclives a la atracción de ramas largas y otros estudios que intentan evitar el error sistemático han dado la siguiente filogenia.[14][15][16][17]
| Protostomia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Un árbol filogenético alternativo publicado en el libro "The Invertebrate Tree Of Life" (2020) es el propuesto por Gonzalo Giribet. Los nodos marcados son defendidos por el autor:[18]
| Árbol filogenético de Protostomia, según Giribet. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Referencias
- Grobben, Karl (1908). «Die Systematische Einteilung des Tierreiches». Zoologisch-Botanischen Gesellschaft, Wien 58: 491-511.
- Arendt, D.; Technau, U.; Wittbrodt, J. (2001). «Evolution of the bilaterian larval foregut». Nature 409 (6816): 81-85. PMID 11343117. doi:10.1038/35051075.
- Hatschek, Berthold (1888). Lehrbuch der Zoologie, 1. Lieferung. Gustav Fisher, Jena.
- Nielsen, Claus (2012). Animal Evolution: Interrelationships of the Living Phyla. Oxford University Press, Oxford.
- Martín-Durán, José M.; Janssen, Ralf; Wennberg, Sofia; Budd, Graham E.; Hejnol, Andreas. «Deuterostomic Development in the Protostome Priapulus caudatus». Current Biology 22 (22): 2161-2166. doi:10.1016/j.cub.2012.09.037.
- Richard C. Brusca, Wendy Moore, Stephen M. Shuster (2016). Invertebrates. Sinauer Associates, Inc. Publishers Sunderland, Massachusetts USA. ISBN 9781605353753.
- Wilson, Edmund B. (1892). «The cell lineage of Nereis». Journal of Morphology 6 (3): 361-480. doi:10.1002/jmor.1050060301.
- Hatschek, Berthold (1891). «Lehrbuch der Zoologie, 3. Lieferung». Gustav Fischer, Jena.
- Nielsen, Claus (2004). «Trochophora Larvae: Cell-Lineages, Ciliary Bands, and Body Regions. 1. Annelida and Mollusca». Journal of Experimental Zoology (Molecular and Developmental Evolution) 302 (1): 35-68. doi:10.1002/jez.b.20001.
- Nielsen, Claus (2005). «Trochophora Larvae: Cell-Lineages, Ciliary Bands and Body Regions. 2. Other Groups and General Discussion». Journal of Experimental Zoology (Molecular and Developmental Evolution) 304 (5): 401-447. doi:10.1002/jez.b.21050.
- Nielsen, Claus (2012). «How to make a protostome». Invertebrate Systematics 26 (1): 25-40. doi:10.1071/IS11041.
- Willmer, Pat (1990). Invertebrate Relationships: Patterns in Animal Evolution. Cambridge University Press, Cambridge.
- Marlétaz, Ferdinand; Peijnenburg, Katja T. C. A.; Goto, Taichiro; Satoh, Noriyuki; Rokhsar, Daniel S. (2019). «A new spiralian phylogeny places the enigmatic arrow worms among gnathiferans». Current Biology 29 (2): 312-318.e3. doi:10.1016/j.cub.2018.11.042.
- Telford, Maximilian J.; Robertson, Helen E.; Schiffer, Philipp H. (18 de junio de 2018). «Orthonectids Are Highly Degenerate Annelid Worms». Current Biology (en inglés) 28 (12): 1970-1974.e3. ISSN 0960-9822. PMID 29861137. doi:10.1016/j.cub.2018.04.088.
- Marlétaz, Ferdinand; Peijnenburg, Katja T. C. A.; Goto, Taichiro; Satoh, Noriyuki; Rokhsar, Daniel S. (2019). «A new spiralian phylogeny places the enigmatic arrow worms among gnathiferans». Current Biology 29 (2): 312-318.e3. doi:10.1016/j.cub.2018.11.042.
- Lahcen I. Campbell, Omar Rota-Stabelli, Gregory D. Edgecombe, Trevor Marchioro, Stuart J. Longhorn, Maximilian J. Telford, Hervé Philippe, Lorena Rebecchi, Kevin J. Peterson, and Davide Pisani (2011). MicroRNAs and phylogenomics resolve the relationships of Tardigrada and suggest that velvet worms are the sister group of Arthropoda. PNAS. https://doi.org/10.1073/pnas.1105499108.
- Marlétaz, Ferdinand (17 de junio de 2019). «Zoology: Worming into the Origin of Bilaterians». Current Biology (en inglés) 29 (12): R577-R579. ISSN 0960-9822. PMID 31211978. doi:10.1016/j.cub.2019.05.006.
- Giribet et Edgecombe.The Invertebrate Tree Of Life. Princeton University Press, 2020. ISBN 06911702589780691170251. pag 21.
Enlaces externos
 Wikispecies tiene un artículo sobre Protostomia.
Wikispecies tiene un artículo sobre Protostomia.
| Control de autoridades |
|
|---|
 Datos: Q5171
Datos: Q5171 Multimedia: Protostomia / Q5171
Multimedia: Protostomia / Q5171- Especies: Protostomia
На других языках
- [es] Protostomia
[ru] Первичноротые
Первичноро́тые[1] (лат. Protostomia) — таксон многоклеточных животных из группы двусторонне-симметричных (Bilateria). В период зародышевого развития на месте их первичного рта (бластопора) образуется рот или, при щелевидном замыкании бластопора, рот и анальное отверстие. Этим они отличаются от вторичноротых, у которых на месте бластопора образуется анальное отверстие, а ротовое возникает позже в другом месте.Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии