bio.wikisort.org - Протисты
Амёба обыкновенная (лат. Amoeba proteus), или амёба протей (корненожка) — относительно крупный (0,2—0,5 мм)[1] амёбоидный организм, представитель класса Lobosea. Полиподиальная форма, характеризуется наличием многочисленных (до 10 и более) псевдоподий — лобоподий, цилиндрических выростов с внутренними токами цитоплазмы.
| Амёба обыкновенная | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Эукариоты Клада: Амёбозои Тип: Tubulinea Класс: Elardia Отряд: Amoebida Семейство: Амёбовые Род: Амёбы Виды: Амёба обыкновенная |
||||||||||
| Международное научное название | ||||||||||
| Amoeba proteus (Pal.) | ||||||||||
| ||||||||||
Строение амёбы
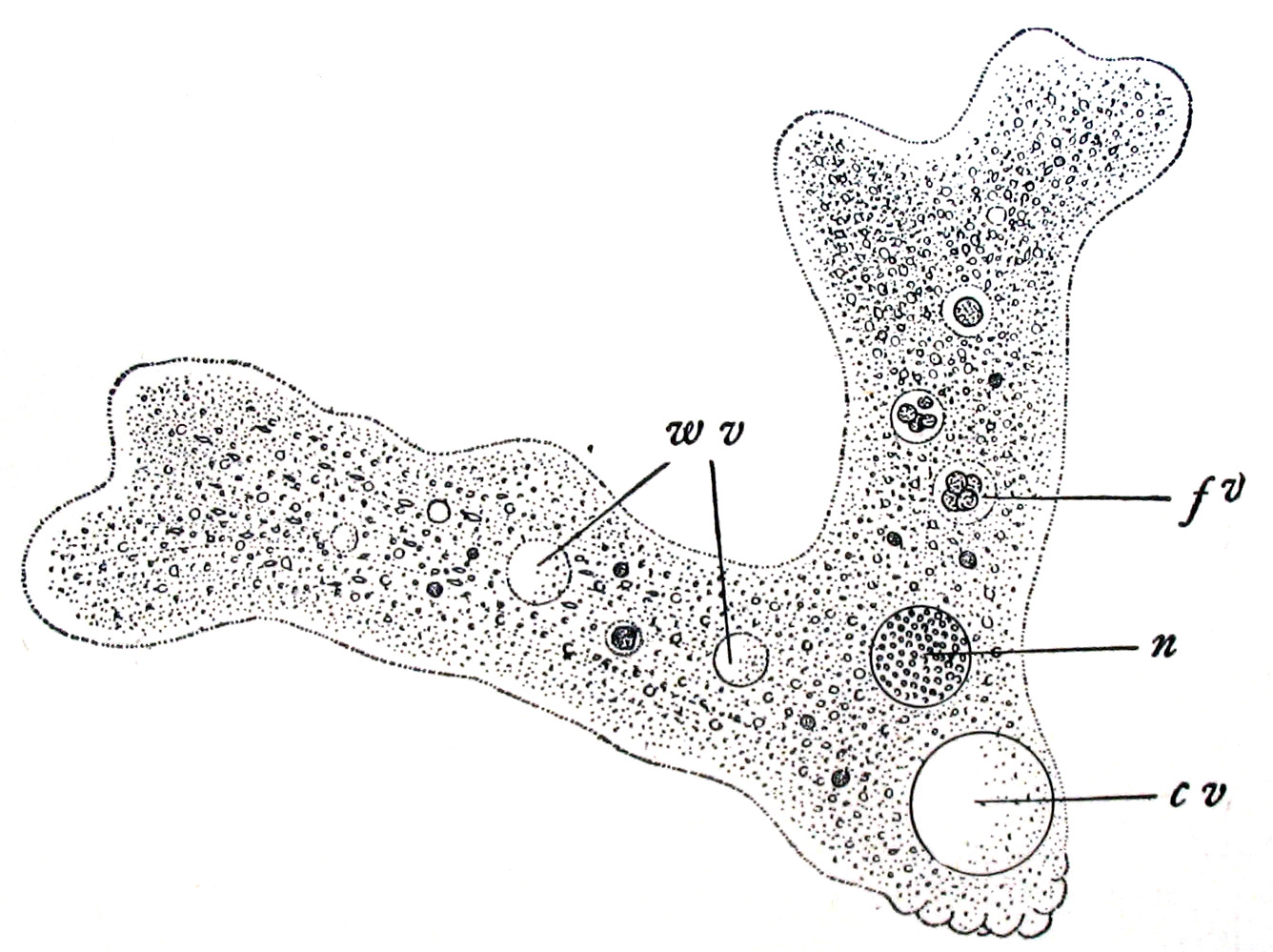
Покров амёбы A. proteus представлен только цитоплазматической мембраной. Вследствие отсутствия твёрдых оболочек клетка имеет непостоянную форму и образует цитоплазматические выросты — псевдоподии (или ложноножки). Цитоплазма клетки дифференцирована на более светлую гелеобразную наружную часть гиалоплазму (эктоплазму), и более темную золеобразную гранулоплазму (эндоплазму), названную так из-за большого содержания различных включений и органелл. Среди клеточных органелл можно выделить одно ядро, одну сократительную вакуоль и множество пищеварительных вакуолей, а также гранул запасных веществ (различных полисахаридов, липидных капель, многочисленных кристаллов).
У данного вида имеется довольно сложный цитоскелет. Гиалоплазма пронизана сетью актиновых и миозиновых микрофиламентов — это кортикальный слой, связанный с клеточной мембраной и окружающий всё содержимое клетки (протопласт). Филаменты располагаются в клетке по-разному. У движущейся амёбы на переднем(«гиалиновом колпачке») и заднем (уроиде) концах актин образует очень тонкий слой, в то время как к середине клетки концентрация актиновых филаментов увеличивается. Миозин на переднем конце клетки также образует тонкий слой, который увеличивается к середине, а на заднем конце, в отличие от актина, — достигает максимальной толщины. Также, различается и их ориентация в пространстве. В передней трети тела движущейся амёбы актиновые филаменты располагаются продольно и соединяются специальными мостиками как с мембраной клетки, так и между собой. В заднем же конце актин формирует трёхмерную сеть, в которой залегают толстые филаменты миозина.
Питание
Амёба протей питается путём фагоцитоза, поглощая бактерии, одноклеточные водоросли и мелких протистов. Образование псевдоподий лежит в основе захвата пищи. На поверхности тела амёбы возникает контакт между плазмалеммой и пищевой частицей, в этом участке образуется «пищевая чашечка». Её стенки смыкаются, в эту область (с помощью лизосом) начинают поступать пищеварительные ферменты. Таким образом формируется пищеварительная вакуоль. Далее она переходит в центральную часть клетки, где подхватывается токами цитоплазмы. Вакуоль с не переваренными остатками пищи подходит к поверхности клетки и сливается с мембраной, таким образом выбрасывая наружу содержимое. Кроме фагоцитоза, амёбе свойствен пиноцитоз — заглатывание жидкости. При этом образуются на поверхности клетки впячивания в форме трубочки, по которой поступает внутрь цитоплазмы капелька жидкости. Образующая вакуоль с жидкостью отшнуровывается от трубочки. После всасывания жидкости вакуоль исчезает. Осморегуляция заключается в том, что в клетке периодически образуется пульсирующая сократительная вакуоль — вакуоль, содержащая излишнюю воду и выводящая её наружу[1].
Движение и реакция на раздражение
Тело Амёбы протей образует выступы — ложноножки. Выпуская ложноножки в определённом направлении, амёба протей передвигается со скоростью около 0,2 мм в минуту. Амёба распознаёт разные микроскопические организмы, служащие ей пищей. Она уползает от яркого света, механического раздражения и повышенных концентраций растворённых в воде веществ (например, от кристаллика поваренной соли).
Основная современная теория амёбоидного движения — теория «генерализованного кортикального сокращения» (Гребецки, 1982). В ней постулируется, что трёхмерное сокращение акто-миозинового комплекса, составляющего кортикальный слой клетки, приводит к сжатию эндоплазмы, в результате чего она направляется к переднему концу клетки, где кортекс наиболее тонкий. Туда же приносятся молекулы глобулярного актина (G-актина), который образуется на заднем конце в результате деполимеризации фибриллярного актина (F-актина), входящего в состав кортекса. В результате этого сокращения в эндоплазме создается повышенное давление, которое продавливает цитоплазму сквозь слой микрофиламентов на её переднем конце как сквозь сито. В результате этого мембрана переднего конца клетки отслаивается от кортекса и выпячивается наружу. Также сквозь филаментозное «сито» проходят и молекулы G-актина (в отличие от крупных включений цитоплазмы), которые затем попадают в пространство между цитоскелетом и мембраной в растущую лобоподию. На внутренней поверхности мембраны расположены специальные центры, полимеризующие G-актин обратно в F-актин, который становится основой для формирования нового цитоскелета. Вновь образованный слой филаментов начинает сокращаться, оказывая на цитоплазму давление, в связи с чем её ток направляется назад, — таким образом прекращается рост лобоподии. В это же время происходит деполимеризация отслоившегося ранее слоя кортекса.
Помимо этой теории, стоит упомянуть и несколько гипотез, предшествовавших ей.
- Гипотеза «потока под давлением» Маста. Предполагалось, что сокращение цитоскелета на заднем конце создается избыточное давление, вызывающее движение эндоплазмы в передний конец клетки, где она расплывается по сторонам, достигая гиалиновой шапочки. В кортикальной зоне происходит переход эндоплазмы в эктоплазму (так называемый золь-гель переход). Из-за того, что эти процессы проходят быстро, создается ощущение непрерывного тока цитоплазмы, в результате которого образуется лобоподия.
- Гипотеза Аллена. Похожа на предыдущую, разве что Аллен считал, что сокращения эндоплазмы происходят не на заднем конце, а на переднем. И там сразу же происходит переход из золя в гель, в результате которого новая порция золеобразной эндоплазмы как бы «подтягивается» к переднему концу, вызывая рост лобоподии. В зоне уроида же происходит обратный переход из геля в золь.
- Гипотеза Серавина. Предположил, что у всех амёбоидных клеток может присутствовать одинаковый набор различных механизмов движения, а различия в движении разных видов формируются в результате разной степени участия того или иного механизма в двигательной активности. Таким образом, согласно Серавину, механизмы, описанные Алленом и Мастом могут иметь место одновременно.
Среда обитания
Обитает на дне пресных водоёмов со стоячей водой, особенно в гниющих прудах и болотах, в которых есть много бактерий. Встречаются локомоторные и флотирующие формы. При плохих для амёбы условиях среды — понижении температуры осенью, пересыхании водоёма — амёба округляется, прекращает потребление пищи и образует плотную оболочку — цисту, а при наступлении хороших — выходит из цисты и ведёт обычный образ жизни[1].
Размножение
Только агамное, бинарное деление. Перед делением амёба перестает ползать, у неё исчезают диктиосомы аппарата Гольджи и сократительная вакуоль. Вначале делится ядро путём митоза, потом происходит цитокинез через образование перетяжки, разделяющей клетку на две равные части с одним ядром в каждой. Размножение стимулируется комфортной температурой и хорошим питанием. При таких условиях скорость размножения составляет 0,5—1 деление в сутки[2]. Половой процесс не описан.
Примечания
- §3. Обыкновенная амёба, ее среда обитания, особенности строения и жизнедеятельности // Биология: Животные: Учебник для 7—8 классов средней школы / Б. Е. Быховский, Е. В. Козлова, А. С. Мончадский и другие; Под редакцией М. А. Козлова. — 23-е изд. — М.: Просвещение, 1993. — С. 11—13. — ISBN 5090043884.
- Полянский Ю. И. Амеба протей // Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1987. — Т. 1 : Простейшие. Кишечнополостные. Черви / под ред. Ю. И. Полянского. — С. 43—45. — 576 с. : ил.
Литература
- Тихомиров И. А., Добровольский А. А., Гранович А. И. Малый практикум по зоологии беспозвоночных. Часть 1. — М.-СПб.: Товарищество научных изданий КМК, 2005. — 304 с.+XIV табл.
Ссылки
- Амёбы — статья из «энциклопедии Кругосвет» в «Единой коллекции цифровых образовательных ресурсов».
- Амёба протей Amoeba proteus (видео)
Другой контент может иметь иную лицензию. Перед использованием материалов сайта WikiSort.org внимательно изучите правила лицензирования конкретных элементов наполнения сайта.
WikiSort.org - проект по пересортировке и дополнению контента Википедии